海洋细菌LP621右旋糖苷酶分批发酵动力学研究
2012-10-23房耀维吴文惠王淑军吕明生焦豫良
葛 亮, 房耀维, 吴文惠, 王淑军, 吕明生, 焦豫良, 刘 姝
(1. 上海海洋大学 食品学院, 上海 201306; 2. 淮海工学院 海洋学院, 江苏 连云港市 222005)
海洋细菌LP621右旋糖苷酶分批发酵动力学研究
葛 亮1,2, 房耀维2, 吴文惠1, 王淑军2, 吕明生2, 焦豫良2, 刘 姝2
(1. 上海海洋大学 食品学院, 上海 201306; 2. 淮海工学院 海洋学院, 江苏 连云港市 222005)
对海洋细菌Pseudoalteromonas tetraodonis LP621右旋糖苷酶进行了50L发酵罐分批发酵的代谢特性的研究。结果表明LP621右旋糖苷酶合成和菌体生长呈部分生长偶联型。据分批发酵试验结果采用 Logistic方程、Luedeking-Piret方程建立了描述菌株生长、产酶以及葡萄糖底物消耗分批发酵动力学模型。动力学模型计算值结果与实验值拟合良好, 较好反映了菌株右旋糖苷酶分批发酵过程的动力学特征。
右旋糖苷酶; 海洋细菌; 发酵动力学模型; Logistic方程; Luedeking-Piret方程
右旋糖苷酶(EC 3.2.1.11)可以切断α-1, 6糖苷键,能够有效阻滞唾液糖蛋白和粘性葡聚糖所组成的口腔牙菌斑形成, 对龋病的防治具有重要的意义[1-2]。同时, 右旋糖苷酶可以用于催化水解高分子右旋糖苷合成血浆替代品, 在制糖工业中也可以用来增加糖的回收率, 降低黏度[3]。另外, 它可以增强药物的化学稳定性和生物利用度[4]。
目前国内外文献报道的右旋糖苷酶生产菌株多来为陆生微生物, 存在的主要问题是酶的最适作用温度高(大多在50℃以上), 酶的最适作用pH低(大多pH 5), 从而导致抑制牙菌斑的效果差, 而海洋微生物因其生活的环境温度较低, 其产生的酶在较低温度下仍具有较高的活性。海洋交替假单胞菌(Pseudoalteromonas tetraodonis)LP621的生长温度为4~37℃, 最适生长温度为 25℃; 在 pH 6. 0~11. 0 条件下生长良好。该菌产生的右旋糖苷酶作用温度低(30℃), 耐热性较好, 具有较好的应用前景[5]。
为了更有效地控制发酵生产[6-7], 提高右旋糖苷酶得率, 本文结合摇瓶发酵条件和发酵罐优化的发酵数据, 通过测定细胞浓度、基质消耗、产物浓度等对时间变化的情况, 观察了海洋细菌LP621在50 L发酵罐的生长及产物生成情况, 建立了细胞生长、产物形成和基质消耗的动力学模型。
1 材料与方法
1.1 菌株
交替假单胞菌(Pseudoalteromonas tetraodonis)LP621[5], 由淮海工学院海洋学院保藏。
1.2 培养基
种子培养基(g/100mL): 2216E 培养基, 酵母粉0.1, 蛋白胨0.5, pH8.0, 陈海水配制。
发酵培养基(g/100mL): 玉米淀粉0.61, 豆饼粉0.4,氯化钠4, 磷酸二氢钾0.1, 硫酸镁0.05, pH6.55。
1.3 主要仪器与设备
50 L全自动发酵罐 MCGS, 上海佰伦科技生产; Synergy HT型酶标仪, 美国BioTek公司产品;分光光度计, 752型, 上海分析仪器厂生产;DK-S24型电热恒温水浴锅, 上海精宏实验设备有限公司生产。
1.4 种子液制备
从 LP621斜面上挑菌一环接于 2216E培养基,25℃, 180 r/min, 培养12 h左右, 再以1%(v/v)的接种量接入750 mL 2216E培养基中, 25℃, 180 r/min, 摇瓶培养12 h, 作为种子液。
1.5 发酵罐发酵
在50L发酵罐中装入30 L发酵培养基, 接种量为 2.5%, 温度控制在 25℃, 在 0~4 h控制通气量为200 L/h, 转速为250 r/min, 4~24 h控制通气量为500L/h, 转速350 r/min, 接种前加入终浓度为0.4%的诱导剂右旋糖苷进行诱导培养。每隔2 h取样150 mL进行菌体干质量、残糖和酶活力的测定。文中所有实验设3个重复, 数据均为3个重复的平均值。
1.6 分析方法
1.6.1 菌体干质量测定
取100 mL发酵液, 10 000 r/min离心5 min, 弃去上清液, 用蒸馏水洗涤将沉淀洗涤, 相同条件离心, 弃上清, 重复3次, 再用蒸馏水洗涤后转移到干燥皿, 80℃烘干到恒质量, 称质量, 即为100 mL菌体的干质量, 换算单位为g/L[8]。
1.6.2 残糖测定
用硫酸蒽酮法测定残糖, 即取离心并稀释一定浓度的发酵液1.0 mL于15 mL刻度试管中, 加入蒽酮试剂3 mL迅速浸入冰水浴中, 再用95℃水浴加热6 min, 冷却, 在分光光度仪上测波长625 nm处吸光度, 通过标准曲线计算残糖的量[9]。
1.6.3 酶活力测定
将发酵液10 000 r/min离心5 min, 取上清液作为粗酶液, 将100 µL粗酶液加入到100 µL 1%的右旋糖苷缓冲液中, 在30℃水浴反应15 min, 用3,5-二硝基水杨酸(DNS)法测定还原糖量。酶活力单位定义: 在上述反应条件下, 每分钟催化产 1 µg麦芽糖的酶量为一个活力单位。
2 结果与讨论
2.1 分批发酵实验的特征
海洋细菌LP621菌株在50 L发酵罐上分批发酵的过程曲线如图1所示, 由图1可知, 菌体在接种以后短时间内就进入快速生长期, 在8~20 h间保持较快生长速度, 当发酵到20 h时细菌进入生长稳定期,菌体干质量保持相对稳定; 产酶曲线走势基本接近细胞干质量曲线趋势, 随时间增长, 酶活逐渐升高,当发酵20 h时, 酶活达到最高值44.97 U/mL, 随后显著下降。海洋细菌 LP621产右旋糖苷酶发酵水平与目前国内外报道的细菌相比较高, 但仍然不及部分真菌来源的右旋糖苷酶发酵水平, 如 Stefan等[10]于2007年分离到一株酵母菌Lipomyces starkeyi酶活力可达到83.9 U/mL。然而, LP621产生的右旋糖苷酶凭借发酵周期短, 安全性高, 耐热性好, 最适作用温度接近人体口腔温度等特点, 使得它在人类牙菌斑的抑制的方面有着难以比拟的优势。
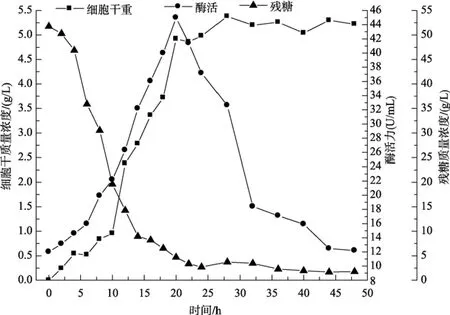
图1 右旋糖苷酶在50L发酵罐分批发酵过程曲线Fig. 1 The batch cultivation curve of LP621 in 50L fermenter
基质的消耗在发酵初期速度较慢, 4 h后下降趋势显著, 在24 h后降到2.70 g/L左右。从菌体生长与右旋糖苷酶合成的关系来看, LP621产生右旋糖苷酶的发酵应属部分生长偶联型。
2.2 发酵动力学模型的建立与拟合分析
2.2.1 菌体生长动力学模型的建立与拟合分析
2.2.1.1 模型建立
由图1可知, LP621的生长曲线接近典型的S型曲线, 采用Logistic方程能够较好地描述菌体的生长规律, 反映出分批发酵过程中, 因菌体浓度的增加对自身生长存在的抵制作用, 并能较好地拟合分批发酵过程的菌体的生长规律。
Logistic 方程:
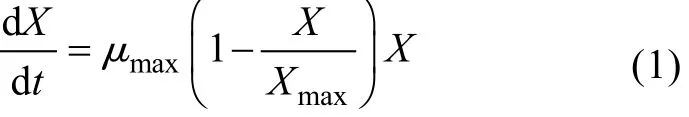
式中,X为细胞质量浓度(g/L);t为时间(h);μ为比生长速率(h-1)
初始条件, 当t=t0时,X=X0, 积分得:
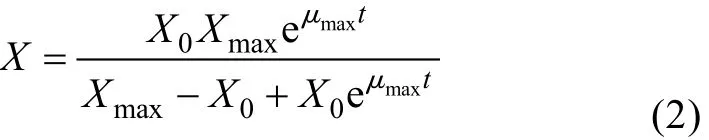
式中,μmax为最大比生长率(h);Xmax为菌体生长上限(g/L)。分批发酵开始时, 菌体浓度很低,X/Xmax项可忽略不计, 式(1)表示菌体呈指数生长; 对数生长结束后, 在稳定期时X=Xmax, 式(1)表示菌体停止生长[11]。
应用Origin8.1软件编程, 进行非线性拟合处理,确定参数估计值, 并求出参数的置信区间, 结果见表1。
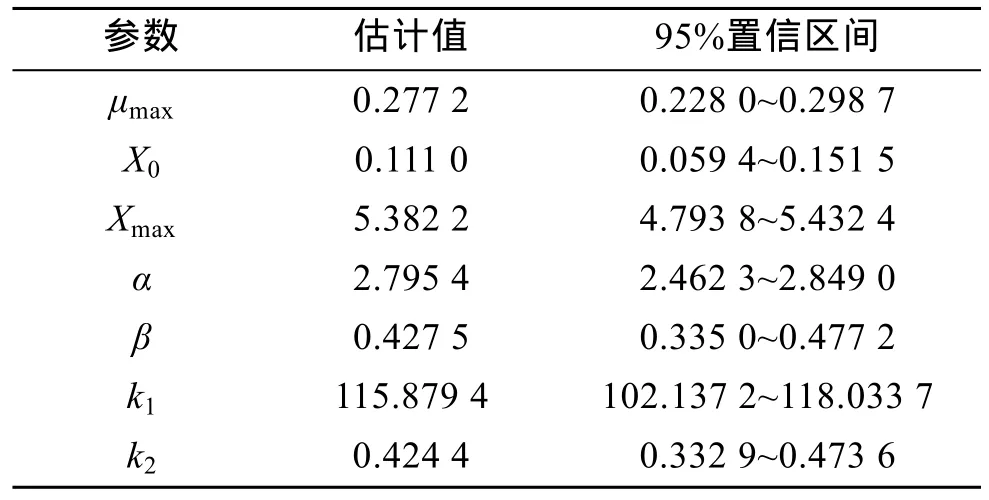
表1 动力学模型参数估计Tab. 1 Parameter values used for testing the model
将参数代入上述方程, 非线性拟合确立的细胞动力学模型为:
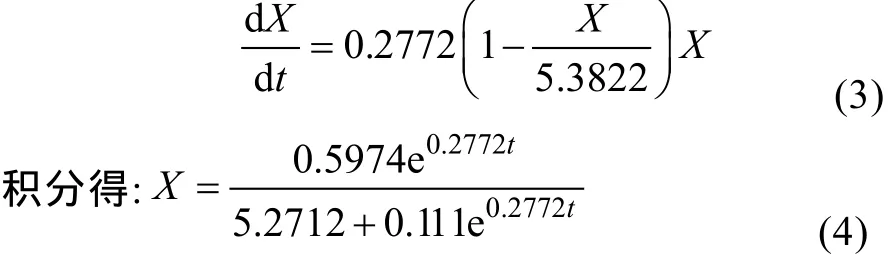
2.2.1.2 拟合分析
由图 2可知, 模型计算细胞干质量的数据与实际测得的数据基本吻合, 实验值与模型值平均相对误差为 7.8%, 这说明 Logistic方程能够较好的模拟50 L发酵罐海洋细菌产右旋糖苷酶过程中的菌体干质量随时间的变化过程, 拟合曲线较为理想。进入稳定期的实验值的点比模型值较高, 可能与营养物质消耗以及细胞利用率有关。
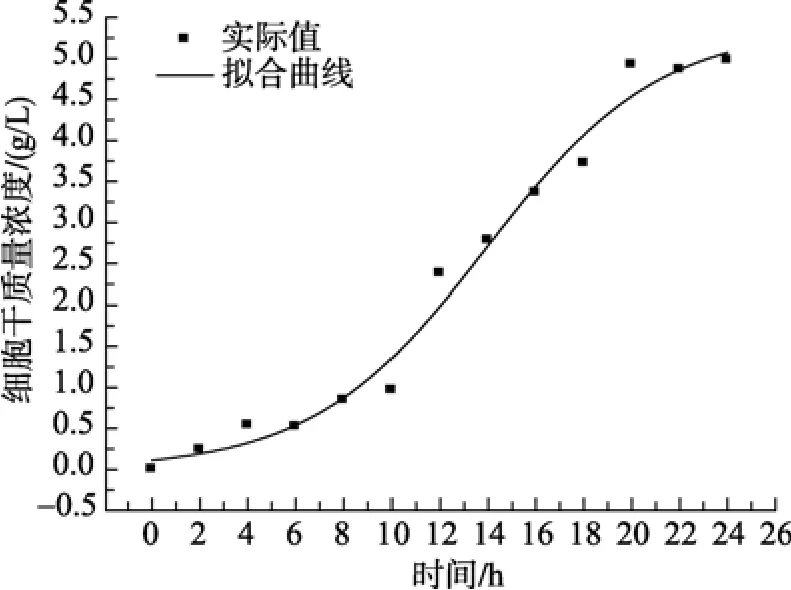
图2 菌体生长的实验值与模型计算值比较Fig. 2 Comparison of experimental data with calculated data of cell growth
2.2.2 产物生成动力学模型的建立与拟合分析
2.2.2.1 模型建立
微生物的产物形成过程非常复杂, 为便于研究,提出了一些定性的描述, 其中 Gaden的观点被广泛接受, Gaden[12]将产物形成分为 3类: 生长偶联型,即只是在菌体生长时才有产物的生成; 部分生长偶联型或称混合型, 即菌体生长阶段有部分产物生成,而大部分产物是在菌体生长稳定期生成的; 非生长偶联型, 即只有在菌体进入稳定期以后产物才开始生成。Luedeking等[13]为此总结了下式描述产物形成与细胞生长的关系。
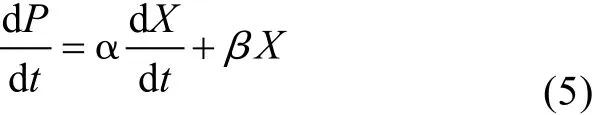
当α>0,β=0 时, 为生长偶联型; 当α>0,β>0 时,为部分生长偶联型; 当α=0,β>0时, 为非生长偶联型。由图1可知, 本实验产物形成与菌株生长的关系属于部分生长偶联型, 故可用 Luedeking-Piret方程来描述其产物生成动力学模型。
应用 Origin8.1软件编程, 将式(3)和(4)代入方程,逐步迭代进行拟合处理, 确定参数估计值(α、β见表1), 并求出参数的置信区间, 代入参数估计值, 得到产物生成动力学模型:
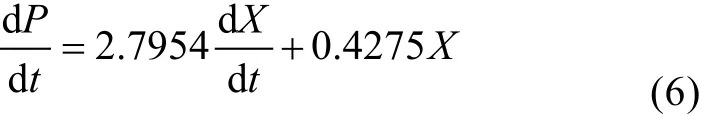
2.2.2.2 拟合分析
2.2.3 基质消耗动力学模型的建立与拟合分析
2.2.3.1 模型建立
发酵过程中, 淀粉消耗主要用于细菌的生长、基本生命活动的维持和代谢产物的生成 3个部分。为方便建模的简化, 可将维持生命活动的基质消耗包括在长菌阶段, 因此右旋糖苷酶的分批发酵过程限制性基质的消耗就可用下式来描述:
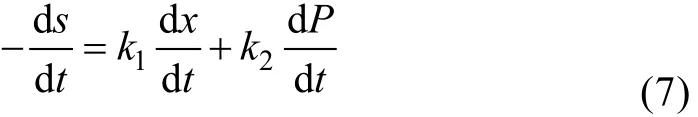
式中,k1表示用于菌体生长的底物消耗常数;k2表示用于产物形成的底物消耗常数。
应用 Origin8.1 软件编程, 将式(3)、(4)和(6)代入方程, 逐步迭代进行拟合处理, 确定参数估计值(k1、k2见表1), 并求出参数的置信区间, 代入参数估计值,得到产物生成动力学模型:

2.2.3.2 拟合分析
由图3可知, 在0~24 h内, 实验值与模型计算值的平均相对误差为 3.4%, 说明所用模型能够较好的描述整个右旋糖苷酶随时间增长的积累过程, 由动力学模型和拟合图形可以看出, 在达到稳定期前右旋糖苷酶形成的类型属于生长偶联型。由图 4可以看出, 该模型能较好地描述海洋细菌 LP621产右旋糖苷酶发酵过程中淀粉的消耗情况, 实验值与模型值的平均相对误差为 7.1%, 说明所拟合曲线能够较好的反映基质消耗与时间的关系, 在接近稳定期时,实验值比模型计算值偏低, 可能是由于底物消耗与菌体生长的不均一所造成的。
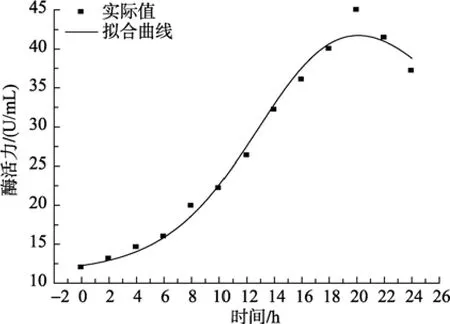
图3 酶活力的实验值与模型计算值比较Fig. 3 Comparison of experimental data with calculated data of amylase activity
3 结论
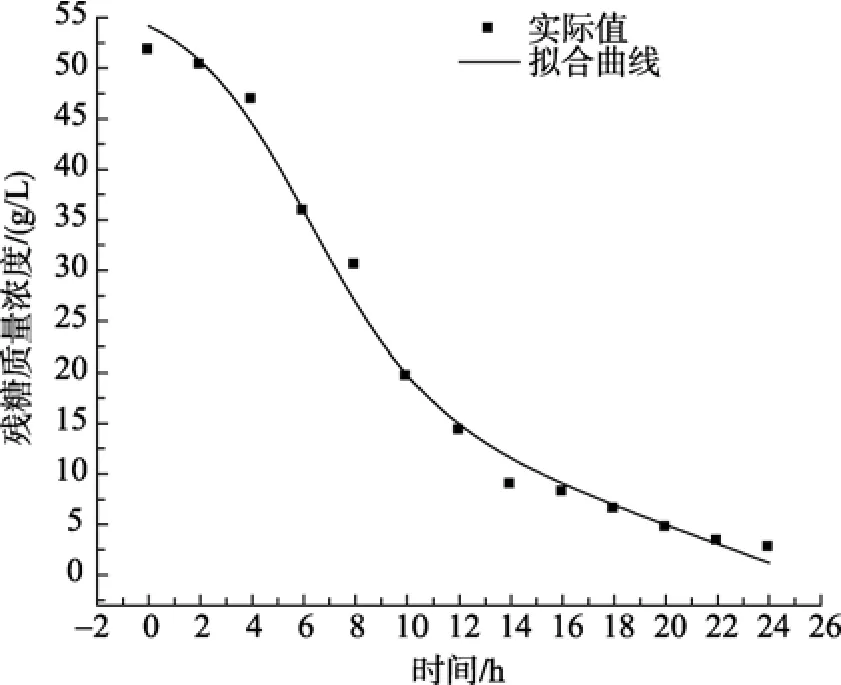
图4 残糖的实验值与模型计算值比较Fig. 4 Comparison of experimental data with calculated data of glucose consumption
本研究在摇瓶发酵条件的基础上, 探讨在 50L发酵罐分批发酵条件和动力学特征。建立了 3个发酵动力学模型, 包括细胞生长动力学模型、产物生成动力学模型和基质消耗动力学模型。这3个发酵动力学模型方程能较好地描述海洋细菌 LP621产右旋糖苷酶发酵的细胞生长动力学、产物生成动力学和基质消耗动力学情况, 解释其动力学规律, 具有很好的适用性, 可为实际发酵过程中细胞生长、产物合成及底物消耗情况的预测与控制提供了理论依据, 为工业化生产设计优化以及经济分析等奠定基础。
[1] Marotta M, Martino A, Rosa A D , et al. Degradation of dental plaque glucans and prevention of glucan formation using commercial enzymes[J]. Process Biochem,2002, 38: 101-108.
[2] Staat R H, Schachtele C F. Characterization of a dextranase produced by an oral strain ofActinomyces israelii[J]. Injection and Immunity, 1975, 12(3): 556-563.
[3] Cuddihy J A, Rauh J S, Mendez F. Dextranase in sugar production: factory experience[J]. A Subsidiary of Associated Chemicals and Services, 1999, 21(7):21-25.
[4] 陈发明, 吴只芬, 金岩, 等. 釉基质蛋白控释微球的研制及其生物学性能的初步研究[J]. 华西口腔医学杂志, 2005, 23(6): 529-533.
[5] 吕明生, 王淑军, 房耀维, 等. 交替假单胞菌 LP621菌株产右旋糖苷酶的培养条件优化[J]. 微生物学杂志, 2010, 30(6): 11-17.
[6] Korz D J, Rinas U, Hellmuth K, et al. Simple fed-batch technique for high cell density cultivation of Escherichia coli[J]. Biotechnol, 1995, 39: 59-65.
[7] 李进, 张伟国. L-异亮氨酸产生菌的选育及发酵条件优化[J]. 江南大学学报, 2005, 43: 7-13.
[8] 韩勇, 黄晓波, 赵良启. Rhizobium sp. N613合成胞外多糖的发酵动力学研究[J]. 化学与生物工程, 2006,23(10): 33-36.
[9] 丁礼琴, 刘力, 徐德生. 蒽酮-硫酸法测定生地黄提取物中总糖的含量[J]. 上海医药, 2008, 29(8): 368-370.
[10] Stefan H. Millson, Ivor Howell Evans, Multiple dextranases from the yeast Lipomyces starkeyi[J]. Antonie van Leeuwenhoek, 2007, 92: 399-404.
[11] 刘永乐, 李忠海, 俞健. 耐酸性 α-淀粉酶发酵动力学的研究[J]. 食品工业科技, 2008, 29(1): 60-62.
[12] Huang J X, Yang J S, Wei Y. Dynamic models for polyhydroxybutyrate fermentation process[J]. Chemical Engineering, 2005, 33(1): 44-47.
[13] Murat E, Ferda M. A kinetic model for actinorbodin production by Streptomyces coelicolor A3[J]. Process Biochemistry, 1999, 34(5): 625-631.
Kinetics analysis for dextranase of Pseudoalteromonas tetraodonis LP621 by liquid fermentation
GE Liang1,2, FANG Yao-wei2, WU Wen-hui1, WANG Shu-jun2, LÜ Ming-sheng2,JIAO Yu-liang2, LIU Shu2
(1. College of Food Science and Technology, Shanghai Ocean University, Shanghai 201306, China; 2. School of Marine Science, Huaihai Institute of Technology, Lianyungang 222005, China)
Dec.,24,2011
dextranase; marine bacteria; fermentation kinetic model; logistic equation; luedeking-Piret equation
The kinetics for dextranase of marine bacteria Pseudoalteromonas tetraodonis LP621 by liquid fermentation was studied. The results showed that the dextranase production was partially associated with the growth of LP621. Based on the result of batch fermentation, the kinetic models for cell growth, dextranase formation, sugar consumption were set up using Logistic equation and Luedeking-Piret equation. The calculated results of models well matched the experimental data. The model equations could well reflect the dextranase fermentation process and kinetic characteristic.
Q815
A
1000-3096(2012)07-0039-05
2011-12-24; 2012-03-22
国家高技术研究发展计划项目(2011AA09070302)
葛亮(1984-), 男, 硕士研究生, 研究方向为海洋天然产物化学, E-mail: frankgl@163.com; 王淑军, 通信作者, 教授, 博士, 研究方向为海洋微生物活性物质, 电话: 0518-85895421, E-mail:shujunwang86@163.com
(本文编辑:梁德海)
