四儿滩地区植物种群的生态位
2012-10-17程中秋张克斌王黎黎赵紫阳
程中秋,张克斌,常 进,王黎黎,王 晓,赵紫阳
(北京林业大学水土保持学院,北京100083)
Grinnell[1]在1917年首次将“生态位”(niche)一词引入生态学领域,并将其定义为“生物在栖息地所占据的空间单元”。随后国外许多学者又提出了多种生态位定义,其中包括Eltonc[2]的“种在其群落中的功能作用和地位”,Hutchinson[3]的“种生存条件的总和”,Odum[4]的“一个种在其群落和生态系统中的地位和状况”等。目前,对于生态位概念的理解趋向于“植物种在多维环境梯度空间的位置”这一表述。生态位理论是生态学中比较成功的理论之一,它已同种间竞争密切联系在一起,而且越来越同资源的利用联系在一起。近十几年国外学者专家对生态位宽度测度、植物种群对资源的分割利用、生态位在不同资源条件下的变化与适应、物种生态位关系和种间竞争与共存的联系等进行了深入研究[5-7]。国内对生态位的测度方法以及生态位的应用研究已做了不少工作[8-13]。
目前,各种植被群落的生态位研究主要集中在高山草甸[14]、沙地植被[15]以及森林优势种群[16]等方面,而关于半干旱区植物种群特别是天然湿地种群的生态位方面研究则相对较少。湿地是由水陆相互作用而形成的自然综合体,是自然界最富生物多样性的生态景观和人类最重要的生存环境之一[17],也被称为“大地之肾”[18]。此前,李瑞等[19]对四儿滩湿地植被进行了较为丰富的研究,但大都从生物多样性和群落空间分布的角度出发,基于生态位理论的研究很少。因此,本研究以四儿滩湿地的植物群落为对象,从生态位角度出发,深入了解湿地植被的特征,以期为半干旱区天然湿地的植被保护和生态恢复提供参考方法。
1 研究区自然概况
盐池县位于宁夏回族自治区东部,地理坐标为37°04′~38°10′N,106°30′~107°41′E。盐池县北与毛乌素沙漠相连,南靠黄土高原,在地理位置上属于一个典型的过渡地带,即:地形上自南向北是从黄土高原向鄂尔多斯台地(沙地)过渡地带,在气候上是从半干旱区向干旱区的过渡地带,在植被上是从干草原向荒漠的过渡地带,在资源利用上是从农区向牧区的过渡地带。这种地理上的过渡性造成了该县自然条件资源的多样性和脆弱性特点。全县地势南高北低,海拔在1 295~1 951 m,南北明显分为黄土丘陵和鄂尔多斯缓坡丘陵两大地貌单元。该县属于典型中温带大陆性气候,年均气温度为8.1℃,极端最高温度为34.9℃,极端最低温度为-24.2℃,年均无霜期为165 d;年降水量仅250~350 mm,且从南向北,从东南向西北递减。土壤类型以灰钙土为主,其次是黑垆土和风沙土,此外有黄土,少量的盐土、白浆土等。植被类型有灌丛、草原、草甸、沙地植被和荒漠植被。其中灌丛、草原、沙地植被数量较大,分布也广。
研究区(图1)位于盐池县城以南8 km处的四儿滩湿地,是哈巴湖国家级自然保护较大的湿地之一。该湿地类型属于沼泽湿地,季节性积水特征明显,水面面积受降水量影响较大。由于湿地盐碱化程度较高,湿地主要生长耐湿、耐盐碱植物,而且主要生长在湿地边缘,如芦苇、盐爪爪、白刺等;草原生态系统由于水分条件较差,其植物类型主要是旱生和中旱生植物,如老瓜头、苦豆子、猪毛蒿等;交错带由于土壤水分条件较好,植物种类丰富,植被类型包括了所有湿地植物和草原植物,同时还有交错带的特有植物,如假芸香、紫菀、香青兰等。
图1 项目区地理位置Fig.1 Location of experimental area
2 研究方法
2.1 样地布设及取样 按照典型性、代表性和科学性的原则,选择有代表性地段,采用GPS定位,设置固定样地进行定位监测。选择每年的7-9月(植物生长季节)进行外业调查。2010年的调查期为7月中旬-8月中旬,调查内容包括植物名称、多度、盖度、高度、地上部分生物量等。调查方法采用样线法,以湿地为中心向东、南、西、北辐射取4条样线,样线长度400 m,跨度包括典型湿地植被带(盐碱滩)、交错带和干旱草原带。按照东样线、西样线、南样线、北样线的顺序分别设置样方。样方设置时考虑到湿地中心30 m范围内几乎为纯盐爪爪群落,为更好地体现不同样线的物种变化情况,本研究排除湿地中心30 m的纯盐爪爪群落,每条样线分别设置15个样方,4条样线共计取样60个,样方大小为1 m×1 m。具体样地布设见图2。
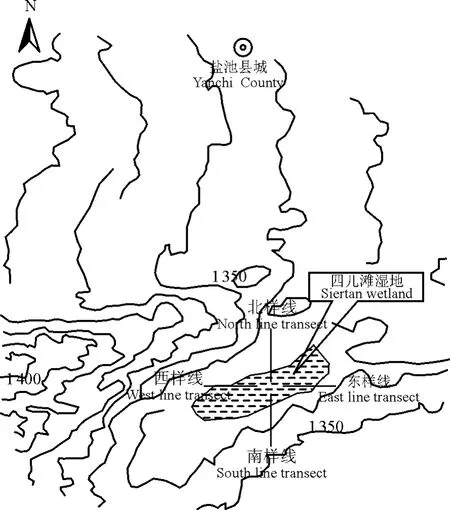
图2 项目区样线布设Fig.2 Sample line layout in experimental area
2.2 数据处理方法 重要值确定各群落的主要成分,以区分不同群落。
重要值[20]=(相对多度+相对频度+相对盖度+相对高度+相对生物量)/5。
生态位宽度采用Levins生态位宽度(Bi)计算公式[21]:
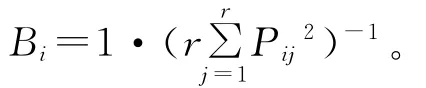
生态位重叠度采用Pianka生态位重叠指数(Oik)[22]计算:
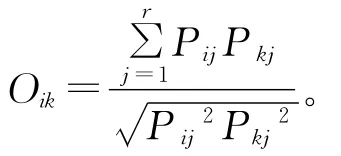
式中,Pij是物种i在第j资源位上的重要值占它所有全部资源位上重要值的比例,r为样方数,Oik为物种i与物种k的重叠度指数。
所有数据采用EXCEL 2007和DPS软件进行计算和分析。
3 结果
3.1 生态位宽度 在2010年调查的60个样方中,四儿滩湿地共出现植物45种(表1),可见湿地生态系统具有较高的物种丰富度。在调查的4条样线中,南样线植物种最多,出现了34种,东样线植物种类最少,共25种,西样线和北样线居中,分别为33种和29种,但各样线植物种数总体相差并不大。
东样线植物种群生态位Levins宽度值从大到小依次是沙鞭、芦苇、苦豆子、虫实、雾冰藜、中亚滨藜、刺沙蓬。其中沙鞭、芦苇、苦豆子的生态位宽度值最大,分别是0.581 9、0.454 6、0.412 9,均超过了0.400 0,显然是东样线植物群落的优势种。而阿尔泰狗娃花、草木樨状黄芪、达乌里胡枝子、假芸香、蒲公英、乳浆大戟、紫菀等植物种生态位宽度值很小,都仅在1个样方中出现,说明这些植物在东样线分布极不均匀,对群落生境的要求极为苛刻,适宜其生存的生境较少,是该样线的特化种。南样线的植物种群生态位宽度前6位的植物种是刺沙蓬、苦豆子、丝叶山苦荬、阿尔泰狗娃花、芨芨草、猪毛蒿,它们是南样线的优势种。其主要优势种的生态位宽度值较为平均,最大的刺沙蓬生态位宽度仅为0.508 7,最小的猪毛蒿其生态位宽度也达到了0.230 5。南样线的植物特化种或伴生种包括糙隐子草、叉枝鸦葱、雾冰藜、西伯利亚蓼等,共16种,均只在调查的15个样方中出现1次。西样线的主要优势种是刺沙蓬、阿尔泰狗娃花、沙鞭、猪毛蒿、碱地风毛菊、西伯利亚蓼,其宽度值均在0.300 0以上,同东样线和南样线相同,其主要优势种的生态位宽度分布较为平均,物种彼此间的生态位宽度值相差不太明显。多裂骆驼蓬、鹅绒藤、狗尾草、赖草等8种植物,均只在样方中出现过1次,是该样线的特化种。北样线植物的生态位宽度值由大到小分别是刺沙蓬、赖草、阿尔泰狗娃花、披针叶黄华、虫实、猪毛蒿。由于生态位宽度值较为接近,宽度值前6位的植物均可以看作该样线的优势种。同样,草地风毛菊、草木樨状黄芪、黑沙蒿等6种植物,均只在样方中出现了1次。
通过对不同样线植物宽度值的统计结果的比较发现,同在四儿滩湿地周边,不同样线的主要优势种各不相同。除少数植物如刺沙蓬在4条样线中均具有较高的生态位宽度外,其他植物很难同时在4条样线中占据主要优势地位。如东样线中生态位宽度值很小的阿尔泰狗娃花,在南、西、北3条样线中却拥有较高的生态位宽度。东、西2条样线中生态位宽度值较大的沙鞭,在南、北2条样线中生态位宽度值却很低。由此可见,在湿地生态系统中,植物种群呈聚集分布的特点。
比较分析4条样线植物种群宽度的平均值可以看出,西样线和北样线的生态位宽度平均值较大,均超过了0.130 0,其中又以西样线为最大,东样线和南样线的生态位宽度平均值较小,都在0.114 0以下。这反映出西、北2条样线的植被生长状况较好,而东、南样线的植被则相对较差。
统计植物种在4条样线总的生态位宽度发现,总宽度值较大的植物种依次是刺沙蓬、阿尔泰狗娃花、沙鞭、苦豆子。所以在四儿滩湿地400 m范围内(排除湿地中心的纯盐爪爪群落),刺沙蓬、阿尔泰狗娃花、沙鞭、苦豆子是该区域的主要优势种,对该生境有较好的适应能力。蒲公英、紫菀、虎尾草、角蒿、香青兰、沙葱、黑沙蒿等在所调查的60个样方中只出现1次,显然是该区的特化种,对该生境的适应性极差。
3.2 生态位重叠度 东样线生态位重叠度最大的是苦豆子和丝叶山苦荬,其重叠度达到0.835,其次是虫实和雾冰藜、多裂骆驼蓬和披针叶黄华、苦豆子和披针叶黄华、沙鞭和芦苇,其重叠度分别是0.828、0.824、0.720、0.715(表2)。作为东样线生态位宽度值最大的沙鞭和芦苇,除其两者之间以及芦苇和赖草的重叠度较大外,它们与其他植物种的重叠度值均在较低水平上,均不足0.400。这充分表明,四儿滩湿地东样线生态位宽度大的物种,与其他物种的重叠度指数不一定大,这一现象从另外一个角度说明了湿地生态系统资源环境存在高度的空间异质性。
南样线生态位重叠度最大的是猪毛蒿和牻牛儿苗,达到0.955(表3),可见二者之间对环境资源具有
相似的生态学要求,具有很强的竞争关系。而该样线的优势种刺沙蓬和芨芨草之间的生态位重叠值为0,说明二者之间生态学特性相差极大。芨芨草是高大多年生密丛禾草,喜生于地下水埋深1.5 m左右的盐碱滩沙质土壤上,在低洼河谷、干河床、湖边、河岸等地分布广泛。刺沙蓬通常生于海拔280~1 400 m的平原盐生荒漠、阿魏蒿荒漠、洪积扇砾质荒漠的小沙堆及河漫滩沙地,对水分的需求相对较小。
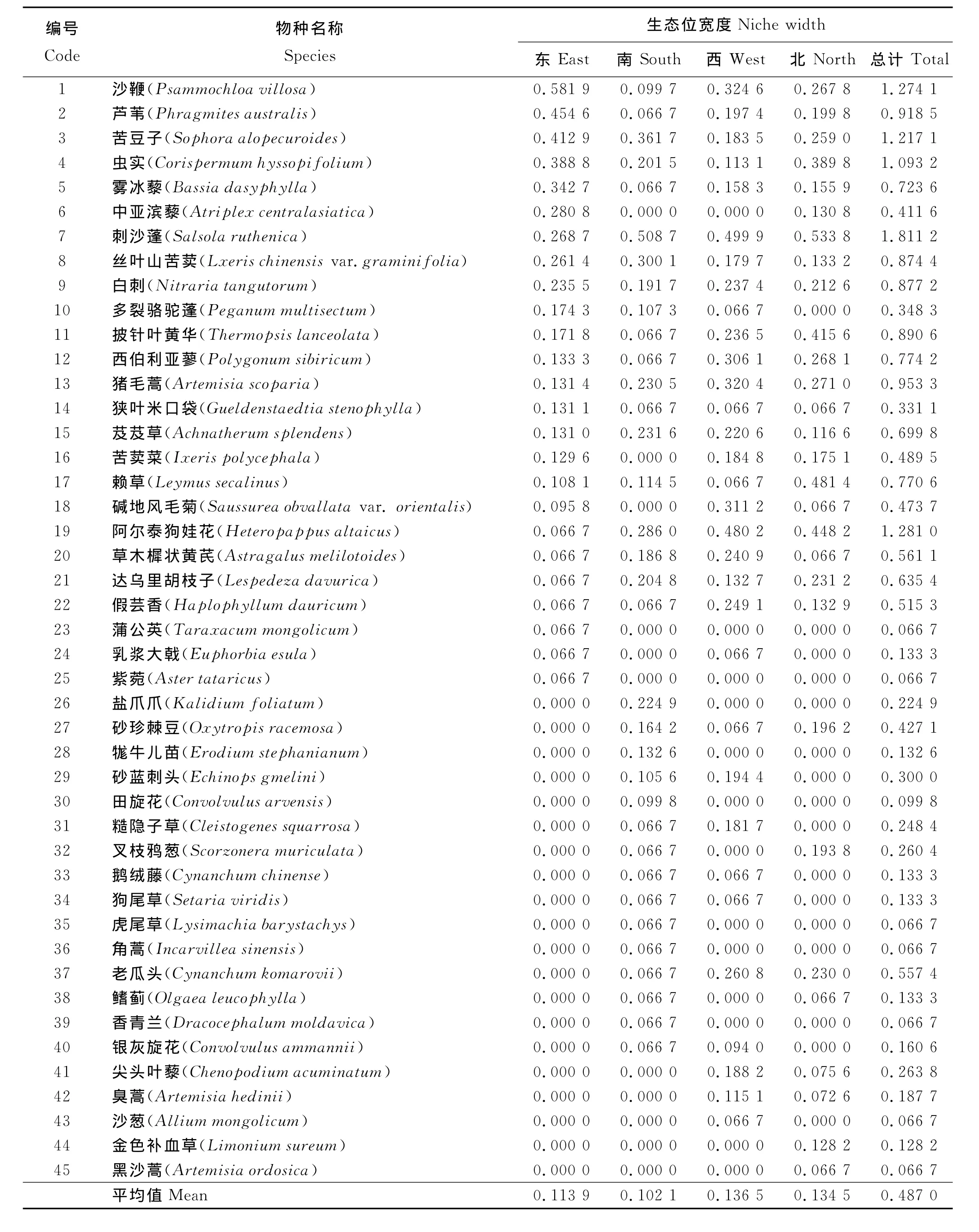
表1 四儿滩湿地植物生态位宽度Table 1 Plant niche width in Siertan wetland

表2 四儿滩湿地东样线生态位重叠度Table 2 Plant niche overlapping of east sampling line in Siertan wetland
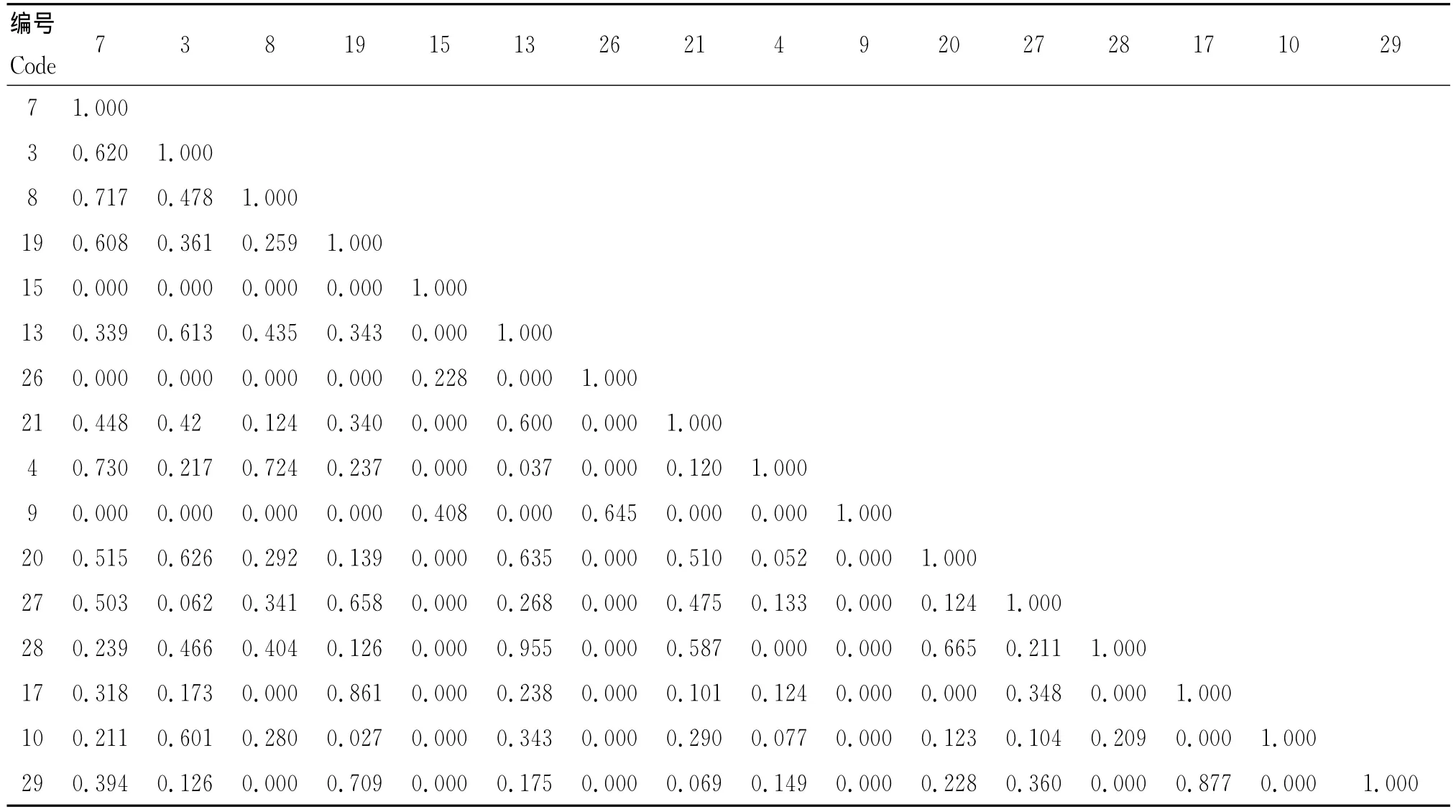
表3 四儿滩湿地南样线生态位重叠度Table 3 Plant niche overlapping of south sampling line in Siertan wetland

?

?
西样线生态位最大重叠度值同样没有发生在宽度值较大的优势种之间,披针叶黄华和砂蓝刺头的重叠度最大,达到0.904(表4)。当然,作为该样线宽度值最大的刺沙蓬和阿尔泰狗娃花也有较高的重叠值,达到0.801,而且之间生态位特性接近,均具有较强的耐旱性。
北样线统计结果更加明显地反映出,生态位宽度大的物种,与其他物种之间的重叠度指数不一定大,除阿尔泰狗娃花和猪毛蒿的重叠度达到0.815外,其他6个重叠度在0.800以上的均出现在宽度值较小的物种之间(表5),这充分说明了湿地生态系统资源环境的空间异质性特点。
4 讨论与结论
生态位宽度反映植物对其生长区域资源环境的利用程度,宽度值越大表明其对各种资源的利用越充分,对当前环境的适应能力越强,在群落中往往处于较高地位,也就是群落的优势种。本研究通过对四儿滩湿地4条不同样线的植物生态位宽度的统计发现,湿地周边不同样线的主要优势种各不相同,这在一定程度上表明湿地生态系统中植物种群呈聚集分布的特点。
生态位重叠度反映物种之间对资源利用的相似程度和竞争关系。生态位重叠度越高意味着物种之间对资源的利用情况越接近,即生态位特性越相似,越可能存在着激烈的竞争[23]。此前多数研究表明[24-25],生态位宽度较大的物种之间存在较高的生态位重叠度。但是四儿滩湿地的生态位重叠度统计结果显示,生态位宽度大的物种与其他物种的重叠度指数不一定大,二者之间并不存在直接的线性关系,这一现象从另外一个角度说明了湿地生态系统资源环境存在高度的空间异质性。
湿地作为西北半荒漠地区十分宝贵的资源,对于维持小区域生态平衡具有重要作用。目前由于西北地区水资源的短缺,湿地入水量呈逐年下降趋势,湿地范围越来越小,周边植物逐渐朝旱生化方向发展。四儿滩湿地作为宁夏盐池县的一个典型湿地,2008年以来随着上游入水量的减少,四儿滩湿地逐渐减小,在湿地中心400 m范围内旱生和强旱生植物逐渐成为该区域的优势种。2010年生态位宽度的调查结果显示,耐旱性较强的刺沙蓬和阿尔泰狗娃花成为四儿滩湿地的主要优势种。因此,保护湿地,使上游水源得到有效补充,对维护半荒漠地区湿地及其周边生态系统的平衡具有重要意义。
[1] Grinnell J.The niche-relationships of the California Thrasher[J].The American Ornithologists’Union,1917,34:427-433.
[2] Elton C S.Animal Ecology[M].London:Sidgwick and Jackson,1927.
[3] Hutchinson G E.Concluding remarks[J].Cold Spring Harbor Symposia on Quantitative Biology,1957,22:415-427.
[4] Odum E P.Fundamentals of Ecology[M].Philadelphia:Saunders,1959.
[5] Arbams P A.Alternative models of character displacement and niche shift 1.Adaptive shifts in resource use when there is competition for nutritionally nonsubstitutable resources[J].Evolution,1987,41(3):651-661.
[6] Berendse F.Inter-specific competition and niche differentiation betweenPlantagolanceolataandAnthoxanthumodoratumin a natural hayfield[J].Journal of Ecology,1983,71:379-390.
[7] Odum E P.Basic Ecology[M].New York:CBS College Publishing,1982:401-407.
[8] 王刚,赵松岭,张鹏云,等.关于生态位定义的探讨及生态位重叠计测公式改进的研究[J].生态学报,1984,4(2):119-127.
[9] 刘金福,洪伟.格氏栲群落生态学研究——格氏栲林主要种群生态位的研究[J].生态学报,1999,19(3):347-352.
[10] 闫美芳,上官铁梁,张金屯,等.五台山蓝花棘砭群落优势种群生态位研究[J].草业学报,2006,15(2):60-67.
[11] 康永祥,岳军伟,雷瑞德,等.陕北黄龙山辽东栎群落优势种群生态位研究[J].西北植物学报,2008,28(3):574-581.
[12] 刘春生,刘鹏,张志祥,等.九龙山濒危植物南方铁杉的生态位研究[J].武汉植物学研究,2009,27(1):55-61.
[13] 张金屯.数量生态学[M].北京:科学出版社,2004.
[14] 李斌,李素清,张金屯,等.云顶山亚高山草甸优势种群生态位研究[J].草地学报,2010,19(1):6-13.
[15] 张继义,赵哈林,张铜会,等.科尔沁沙地植物群落恢复演替系列种群生态位动态特征[J].生态学报,2003,23(12):2741-2746.
[16] 苏志尧,吴大荣,陈北光,等.粤北天然林优势种群生态位研究[J].应用生态学报,2003,14(1):54-59.
[17] 陈宜瑜.中国湿地研究[M].长春:吉林科学技术出版社,1995:1-7.
[18] Mitsch W J,Gosselink J G.Wetlands[M].New York:Van Nostrand Reinhold Company Inc,1986:89-125.
[19] 李瑞,张克斌,王百田,等.湿地——干草原生态系统植物物种多样性研究[J].北京林业大学学报,2006,28(5):12-17.
[20] 郑翠玲,曹子龙,赵廷宁,等.浑善达克沙地南缘农牧交错带弃耕地植被的演替规律[J].中国水土保持科学,2005,3(1):72-76.
[21] 魏志琴,李旭光,郝云庆.珍稀濒危植物群落主要种群生态位特征研究[J].西南农业大学学报:自然科学版,2004,26(1):1-4.
[22] 毕润成,尹文兵,王艳妮.山西南部脱皮榆种群生态位的研究[J].西北植物学报,2003,23(7):1266-1271.
[23] Silvertown J W.Plants in limestone pavements:tests of species interactionand niche separation[J].Journal of Ecology,1983,71:819-828.
[24] 苏志尧,吴大荣,陈北光.粤北天然林优势种群生态位研究[J].应用生态学报,2003,14(1):101-105.
[25] 董全民,赵新全,马玉寿,等.高寒小嵩草草甸暖季草场主要植物种群的生态位[J].生态学杂志,2006,25(11):1323-1327.
