马铃薯甲虫自然种群抗寒能力测定
2012-08-27张云慧吐尔逊程登发
张云慧, 张 智, 何 江, 吐尔逊, 程登发*
(1.中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193;2.新疆农业科学院植物保护研究所,乌鲁木齐 830000)
马铃薯甲虫自然种群抗寒能力测定
张云慧1, 张 智1, 何 江2, 吐尔逊2, 程登发1*
(1.中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193;2.新疆农业科学院植物保护研究所,乌鲁木齐 830000)
为明确马铃薯甲虫自然种群的抗寒性,利用热敏电阻测定仪对马铃薯甲虫不同虫态过冷却点和冰点进行了测定。结果表明:不同虫态过冷却点和冰点之间存在显著性差异,其中以卵和成虫的过冷却点最低,分别为:(-13.73±0.93)℃和(-7.89±0.07)℃,显著低于其他虫态。3龄幼虫的过冷却点和冰点最高,分别为:(-6.21±0.08)℃和(-2.80±0.08)℃;过冷却点由低到高顺序为:卵<成虫<1龄幼虫<蛹<4龄幼虫<2龄幼虫<3龄幼虫;冰点由低到高的顺序为:卵<1龄幼虫<成虫<2龄幼虫<蛹<4龄幼虫<3龄幼虫;同一虫态个体间的过冷却点频次分布出现不同程度的变异,但均服从正态分布。研究结果为制定马铃薯甲虫在我国潜在分布和风险分析提供了科学依据。
马铃薯甲虫; 抗寒性; 过冷却点; 结冰点
昆虫是变温动物,温度对其影响是不言而喻的,尤其低温作为一个重要的因素影响着它们的发生、分布、生殖和扩散[1]。因此,明确昆虫的耐寒性对于其在一个国家和地区的分布,能否向北扩散并建立稳定的种群,以及成功预测预报和根治其危害都起着至关重要的作用[2]。对昆虫耐寒性的研究最早可追溯至18世纪30年代,Bachmetjew发现了昆虫的过冷却点,这一发现为昆虫在低温环境下的适应能力,以及许多有关实践问题的研究开辟了广阔的前景[3]。吴孔明等研究了我国不同地理种群棉铃虫的抗寒性,并首次提出我国棉铃虫越冬的北界,提高了我国棉铃虫预报的准确性,为建立我国的棉铃虫区域性灾变预警技术体系奠定了基础[4]。韩兰芝等通过测定甜菜夜蛾不同地理种群的过冷却点和结冰点,结合田间埋蛹越冬试验和气温分析,得出甜菜夜蛾的常年越冬区范围应在南京以南[5]。一般情况下昆虫的过冷却能力越强,其抗寒能力就越强。因而在许多昆虫耐寒性的研究中,过冷却点成为界定其耐寒性强弱的重要指标之一[6-7]。因此,测定昆虫的过冷却点,对于了解其抗寒能力,预测越冬死亡率和种群变动规律,确定科学防控措施,以及深入研究该虫的低温生态学,都具有理论和生产的双重意义。
马铃薯甲虫[Leptinotarsadecemlineata(Say)]发源地为墨西哥北部的洛基山东麓,该虫分布于欧洲、非洲、亚洲和北美洲的30多个国家和地区,是一种极具毁灭性的检疫性害虫[8]。在我国新疆主要以成虫、幼虫进行为害,给马铃薯生产造成严重的经济损失,一般减产30%~50%,有时高达90%,甚至绝收[9]。1993年,马铃薯甲虫在新疆西部伊犁自治州察布查尔县、塔城市被首次发现,在随后的几年中,马铃薯甲虫逐渐沿着伊犁河谷向东、向北扩散,最东端到达乌鲁木齐的木垒县,目前分布于新疆的36个县市,成为新疆马铃薯生产的重要害虫,并对其他马铃薯主产区形成严重威胁[10]。自马铃薯甲虫传入我国新疆地区,围绕马铃薯甲虫的发生规律和防控技术开展了大量的研究[11-13],但是有关马铃薯甲虫抗寒性研究,目前国内仍属空白。国际方面发表的相关报道也不多,相关研究综述和学位论文中引用的多是俄罗斯研究发表的一篇文献。该研究结果表明滞育的马铃薯甲虫过冷却点在-6.15~-5.7℃[14]。本研究团队自2008年以来,通过对马铃薯甲虫越冬区的调查和地温的收集发现马铃薯甲虫过冷却点结论与我国新疆马铃薯甲虫的实际的越冬区有些出入。因此,有必要弄清我国新疆地区马铃薯甲虫的抗寒性,结合各地气象数据建立马铃薯甲虫在我国的越冬区划,为马铃薯甲虫的监测预警提供基础生物学依据。本研究从马铃薯甲虫的抗寒性入手,测定马铃薯甲虫不同虫态的过冷却点和冰点,为研究马铃薯甲虫在我国潜在分布和风险分析提供科学依据。
1 材料与方法
1.1 供试虫源
供试马铃薯甲虫成虫采自乌鲁木齐市安宁渠新疆农科院试验场“国家现代农业示范区”(87°28′E,43°56′N)实验基地马铃薯试验田,直接从田间随机采集越冬代成虫,卵、幼虫、蛹。测试时间为5月上旬马铃薯甲虫越冬代成虫出土,到6月中旬第1代幼虫化蛹盛期。
1.2 实验仪器
DW-40型低温试验箱(-40~120℃)由浙江上虞市龙翔精密仪器厂生产,降温速度1℃/min;SUN-Ⅱ型智能过冷却点测定仪及配套软件由中国农业科学院植物保护研究所和鹏程电子有限公司合作研发。每处理测定试虫40头,5s1次由昆虫过冷却仪的数据采集器自动采集虫体温度变化后输入计算机,并用相应的软件对数据进行处理,实时绘制虫体温度变化曲线,数据存储为txt格式。当降到一定温度时,昆虫体液开始结冰,此时的温度即为过冷却点,虫体结冰后释放出热量,在温度变化曲线上显示为跳跃峰值,峰值即为结冰点[15]。
1.3 过冷却点和冰点的测试方法
把田间采集的马铃薯甲虫成虫和幼虫放置在宽4.5cm,长6cm的塑料透明胶带上,将过冷却点测定仪的热敏电阻感温探头与虫体充分接触,用胶带把虫体与探头包好,使两者充分接触后置于高低温实验箱中。蛹和卵置于0.8mL的离心管中,将热敏电阻感温探头深入离心管,使探头与虫体充分接触后,用脱脂棉封口置于高低温实验箱中。
1.4 数据分析
计算不同虫期过冷却点和冰点的平均值和标准差,用Duncan多重比较进行差异显著性测定,并采用单样本Kolmogorov-Smirnov检验分析不同虫期过冷却点的频次分布特征。所有数据处理均采用SPSS 16.0(SPSS Inc.,Chicago)进行。
2 结果与分析
2.1 马铃薯甲虫过冷却点测定
马铃薯甲虫各虫态过冷却点存在一定的差异,测定结果见表1,以卵的过冷却点最低为(-13.73±0.93)℃,其次为成虫和1龄幼虫分别为:(-7.89±0.07)℃和(-7.87±0.18)℃,成虫和1龄幼虫之间无显著性差异,但显著低于其他虫态(p<0.05);其次为蛹(-6.90±0.12)℃,显著低于2~4龄幼虫(p<0.05);2~4龄幼虫过冷却点较高,它们之间没有显著性差异(p>0.05),幼虫各龄期平均过冷却点为(-6.66±0.08)℃,幼虫各龄期中以3龄幼虫过冷却点最高(-6.21±0.08)℃。过冷却点由低到高的具体顺序为:卵<成虫<1龄幼虫<蛹<4龄幼虫<2龄幼虫<3龄幼虫。
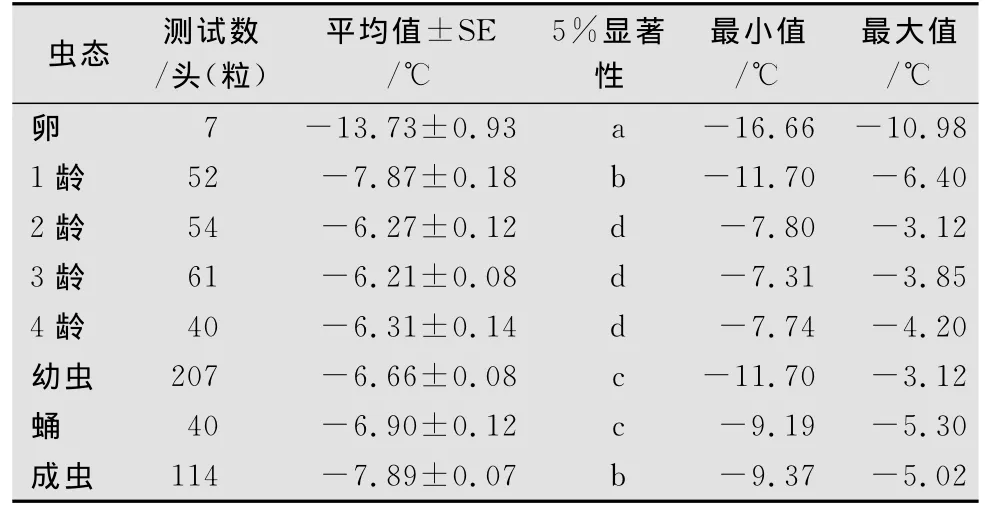
表1 马铃薯甲虫不同虫态过冷却点
2.2 马铃薯甲虫冰点的测定
马铃薯甲虫冰点也存在一定的差异,不同发育阶段的冰点测定结果见表2。冰点由低到高的顺序为:卵<1龄幼虫<成虫<2龄幼虫<蛹<4龄幼虫<3龄幼虫。冰点平均值最低为卵(-12.57±0.94)℃,其次为1龄幼虫和成虫,分别为:(-6.64±0.18)℃和(-6.57±0.79)℃,两者之间没有显著性差异,冰点最高值为3龄幼虫(-2.80±0.08)℃。不同个体之间冰点极限最低值为卵-15.39℃,最高值为4龄幼虫-0.51℃,不同虫态之间冰点存在较明显的差异(p<0.05)。方差分析表明,不同虫态冰点的变化趋势和不同虫态过冷却点的变化趋势基本一致。
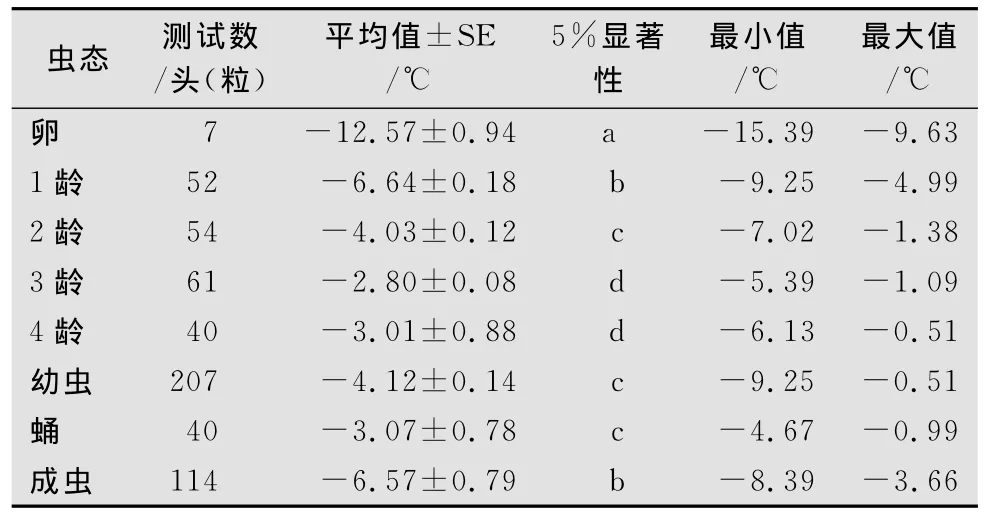
表2 马铃薯甲虫不同虫态冰点
2.3 过冷却点在各虫态的频次分布
马铃薯甲虫各虫态个体的过冷却点出现不同程度的变异(图1)。各虫态个体的过冷却点分布范围具有一定宽度。过冷却点最低值为卵期-16.66℃,最高值出现在2龄幼虫-3.12℃。卵的过冷却点分布范围最宽,最低值与最高值相差5.68℃。3龄幼虫过冷却点分布范围最小,最低和最高值相差3.46℃。具体宽度范围由大到小为:卵>1龄幼虫>2龄幼虫>成虫>蛹>4龄幼虫>3龄幼虫。各虫期的过冷却点频次均服从正态分布(p>0.05)。
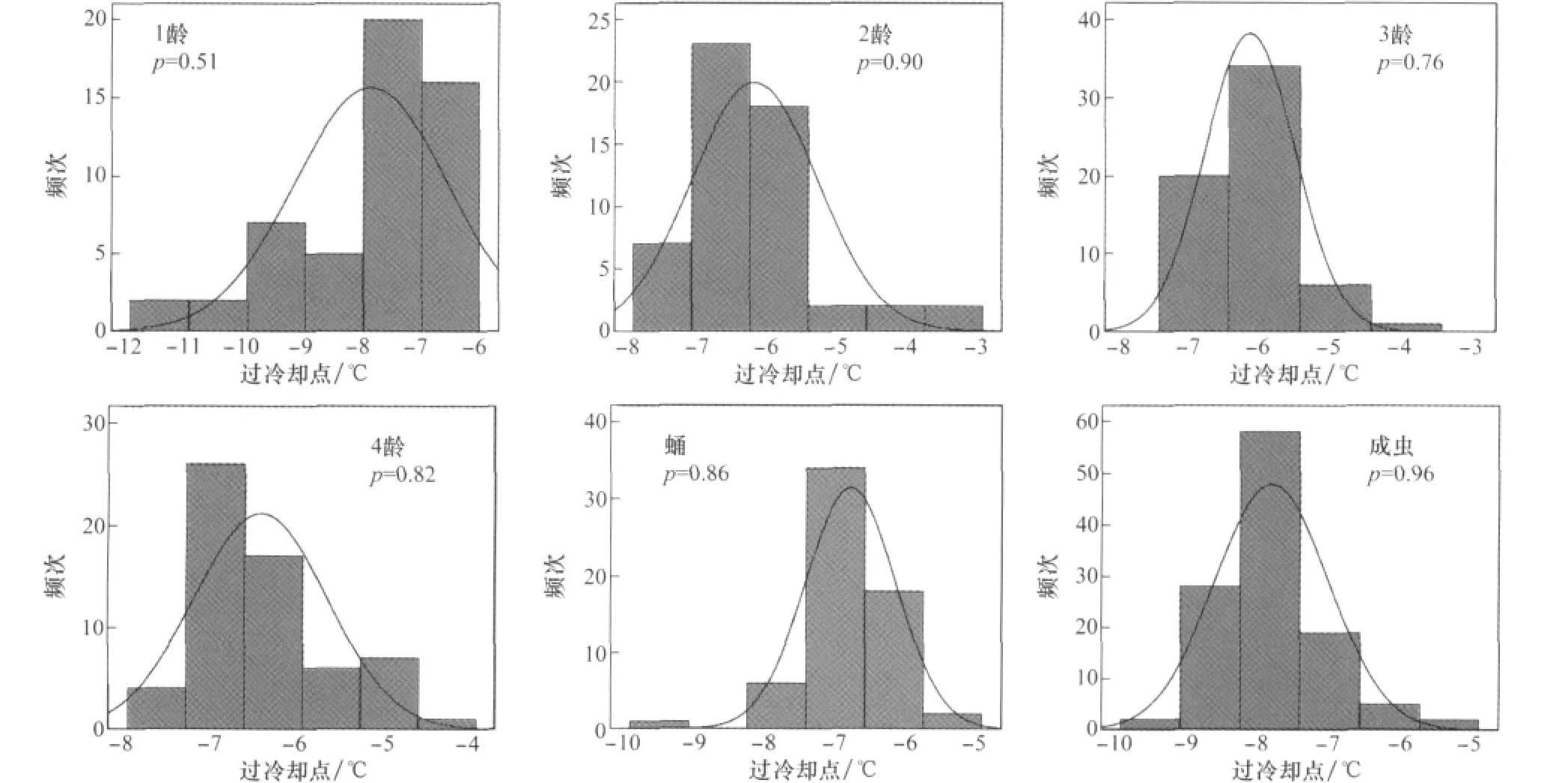
图1 马铃薯甲虫各虫态过冷却点频次分布
3 讨论
入侵物种往往能形成极具优势性的暴发与扩张,很大程度上取决于它强大的竞争以及对恶劣环境的适应能力[2]。例如南美斑潜蝇向北扩散的过程中同时伴随着过冷却点降低的适应机制[16]。早期的研究结果也表明马铃薯甲虫具有极强的生理、生态适应能力,当新入侵地区环境条件不利于其发育时,该虫可通过滞育等一系列生物学机制适应不良环境,在3~5年时间内,种群就能适应新的环境,并表现出与原始种群不同的生理生态学特性[17]。在马铃薯甲虫越冬成虫的抗寒性研究方面不同时间、不同地区过冷却点存在很大差异,例如俄罗斯20世纪70年代的结果表明滞育状态下的马铃薯甲虫过冷却点在(-6.15~-5.7)℃[14],而加拿大专家20世纪90年代研究结果为越冬后马铃薯甲虫平均过冷却点为-8.8℃,土中滞育状态下平均过冷却点可达-11.7℃,从而保证种群向北延伸到加拿大的新不伦瑞克省[18]。本文测定的马铃薯越冬成虫过冷却点为(-7.89±0.07)℃,与俄罗斯和加拿大专家的研究结果也存在一定的差异,充分显示了马铃薯甲虫作为入侵性害虫超强的生理生态适应机制。
另外,对于马铃薯甲虫整个生育期的抗寒性测定结果显示以卵和成虫的过冷却点和冰点最低,其次为1龄幼虫,符合马铃薯甲虫的抗逆性对策。马铃薯甲虫以成虫越冬,在我国新疆地区每年4月下旬越冬代成虫开始出土,成虫产卵始期为5月上旬,第1代幼虫发生始期为5月中旬[10]。根据新疆气象资料4月下旬至5月中旬正是马铃薯甲虫发生区气温最不稳定、常有寒流发生、昼夜温差大的时期,生活在这个时期的越冬代成虫、卵和1龄幼虫通过更高的耐寒性适应环境的胁迫。试验过程中由于卵个体较小,给测试带来了一定的困难,本试验共测试200粒卵,只有7粒得到试验结果。关于卵的耐寒性需要进一步改进实验仪器进行测定,但从7个成功的测试结果显示,卵具有较高的抗寒性,这和另一个入侵害虫扶桑绵粉蚧抗寒对策一致,卵和1龄幼虫具有更强的抗寒性[19]。
本文对马铃薯甲虫的过冷却点测定发现:同一发育阶段的过冷却点有一定的变异,从频次分布图看均符合正态分布,表明过冷却点在同一发育阶段的不同个体之间耐寒性存在差异。Klok和Chown认为这种现象是变温动物的一种适应策略,在遭受突如其来的意外的寒冷情况下,抗逆性强的个体也能存活下来[20]。另外,昆虫的过冷却点在很多种类中不是稳定和集中的,它会因环境、营养、发育阶段和虫体的结构不同而不同,还会受各种环境因素的影响,例如:如气候的季节变化,纬度或海拔的差异等,以及昆虫自身的发育阶段、滞育发生等,这种变化与营养、体内化学物质组成、虫体结构有一定关系[7,21-22]。因此,不少学者将过冷却点整合到种群在地理分布范围扩散方面的研究,探讨昆虫在种群扩散过程中过冷却点变化对种群扩散的生态学意义[4,15,23]。尤其是研究毁灭性检疫害虫种群的耐寒性分化对预测其扩散机会和综合治理该类害虫有十分重要的意义,下一步拟从马铃薯甲虫体内抗寒物质入手,通过全面掌握其耐寒机制,在获得该虫在不同环境因子下发生危害规律等科学数据后,应用CLIMEX等软件进行预测,获得更科学的结果。
[1]景晓红,康乐.昆虫耐寒性的测定与评价方法[J].昆虫知识,2004,40(1):7-10.
[2]许益镌,陆永跃,黄俊,等.红火蚁自然种群耐寒性的研究[J].昆虫学报,2009,52(9):974-983.
[3]Saltrw.Principles of insect cold-hardiness[J].Annual Review of Entomology,1961,6(3):55-74.
[4]吴孔明,郭予元.棉铃虫的抗寒能力[J].生态学报,1997,17(3):298-302.
[5]韩兰芝,翟保平,张孝羲.甜菜夜蛾的抗寒力研究[J].植物保护学报,2005,32(2):169-173.
[6]Renault D,Salin C,Vannier G,et al.Survivalat low temperatures in insects:What is the ecological significance of the supercooling point?[J].CryoLetters,2002,23:217-228.
[7]Leather S R,Walters K F A,Bale J S.The ecology of insect overwintering [M]. Cambridge: Cambridge University Press,1993.
[8]赵建周.国外马铃薯甲虫发生危害与防治概况[J].植物保护,1995,21(4):35-36.
[9]张润志,刘宁,李颖超.危害茄科蔬菜的外来入侵害虫-马铃薯甲虫[J].生命科学,2010,22(11):1118-1121.
[10]郭文超,吐尔逊,许建军,等.马铃薯甲虫识别及其在新疆的分布、传播和危害[J].新疆农业科学,2010,47(5):906-909.
[11]洪波,程登发,吐尔逊,等.新疆地区马铃薯甲虫发育与温度关系的研究[J].植物保护,2009,35(6):60-63.
[12]王春林.外来入侵生物(马铃薯甲虫)的预警研究[D].重庆:重庆大学,2005.
[13]张润志.马铃薯甲虫发生规律和防治技术研究[D].北京:中国科学院研究生院,1996.
[14]Minder I F,Chesnek S I.The dependence of cold resistance in the Colorado potato beetle on the time of the onset of diapause[J].Zoologicheskii Zhurnal,1970,49(6):855-861.
[15]赵琦,张云慧,韩二宾,等.旋幽夜蛾各虫态的过冷却点测定[J].植物保护,2011,37(2):63-66.
[16]陈兵,康乐.南美斑潜蝇地理种群蛹过冷却点随纬度递变及其对种群扩散的意义[J].动物学研究,2003,24(3):168-172.
[17]Ushatinskaya R S,Ivanchik E P.Reversibility of ecologophysiological adaptations in the Colorado potato beetle[J].Zoologicheskii Zhurnal,1982,61(3):358-363.
[18]Gilles B,Warren C.Cold tolerance in the Colorado potato beetle,Leptinotarsadecemlineata(Say)(Coleoptera:Chrysomelidae)[J].The Canadian Entomologist,1996,128(6):1087-1099.
[19]关鑫,陆永跃,曾玲,等.扶桑绵粉蚧的过冷却点和体液结冰点测定[J].环境昆虫学报,2009,31(4):381-384.
[20]Klok C J,Chown S L.Interactions between desiccation resistance,host plant contact and the thermal biology of aleaf-dwelling sub-antarctic caterpillar,Embryonopsishalticella(Lepidoptera:Yponomeutidae)[J].Journal of Insect Physiology,1998,44:615-628.
[21]刘宁,文丽萍,何康来,等.不同地理种群亚洲玉米螟抗寒力研究[J].植物保护学报,2005,32(2):163-168.
[22]张珺,吴孔明,林克剑,等.二化螟温带和亚热带地理种群的滞育特征与抗寒性差异[J].中国农业科学,2005,38(12):2451-2456.
[23]江幸福.甜菜夜蛾抗寒与越冬能力研究[J].生态学报,2011,21(10):1575-1582.
Cold hardiness of natural populations of the colorado potato beetle,Leptinotarsadecemlineata
Zhang Yunhui1, Zhang Zhi1, He Jiang2, Tuerxun2, Cheng Dengfa1
(1.InstituteofPlantProtection,ChineseAcademyofAgriculturalSciences,Beijing100193,China;2.InstituteofPlantProtection,XinjiangAcademyofAgriculturalSciences,Urumqi830000,China)
The supercooling points and freezing points of the colorado potato beetle,Leptinotarsadecemlineataat all stages,were measured using a thermocouple.It showed that the supercooling points and freezing points were significantly different between adults,eggs,larvae and pupae,in which the eggs and adults had stronger cold hardness(-13.73±0.93℃and-7.89±0.07℃)than other stages,and the differences were significant.The highest supercooling point and freezing point of the 3rd-instar larvae were-6.21±0.08℃and-2.80±0.08℃,respectively.The sequence of supercooling points from low to high was egg,adult,the 1st-instar larva,pupa,the 4th-instar larva,the 2nd-instar larva and the 3rd-instar larva.And the sequence of freezing points from low to high was egg,the 1st-instar larva,adult,the 2nd-instar larva,pupa,the 4th-instar larva and the 3rd-instar larva.The supercooling point ofL.decemlineatavaried significantly among individuals,but followed a normal distribution.The results provide a scientific basis for the development of risk analysis and potential distribution ofL.decemlineata.
Leptinotarsadecemlineata; cold hardiness; supercooling point; freezing point
S 435.32
A
10.3969/j.issn.0529-1542.2012.05.012
2012-01-18
2012-03-21
公益性行业(农业)科研专项(201103026-2)
* 通信作者E-mail:dfcheng@ippcaas.cn
