珠江口夏季中、小型浮游动物生态特征研究
2012-08-14吴玲玲朱艾嘉郭娟方宏达蔡伟叙张敬怀
吴玲玲,朱艾嘉,郭娟,方宏达,蔡伟叙,张敬怀
(国家海洋局南海环境监测中心,广东 广州 510300)
浮游动物是河口生态系统食物网中的关键环节,它们下行控制初级生产力,上行被经济鱼类等高级消费者摄食,是河口生态系统结构和生源要素循环中扮演着重要角色。中、小型浮游动物具有繁殖快、生命周期短、产量高、粒度小的特点,其对浮游植物的摄食压力会超过大型浮游动物,对海洋初级生产力的利用大于大型浮游动物(Morales et al,1991)。此外,小型桡足类是仔稚鱼最重要的开口饵料,作为基础饵料比大型桡足类更具特殊意义,中、小型浮游动物数量成为判断经济鱼类资源量的重要依据(王荣等,2002;熊瑛等,2009)。
珠江口位于南海北部,属于典型亚热带河口。珠江口海域内的伶仃洋河口湾是一个喇叭型河口湾,其纵长约50 km,宽度由北向南逐渐增大,虎门口附近宽仅为8 km,内伶仃岛附近宽达30 km。珠江三角洲地区是我国改革开放的先行地区,是我国重要的经济中心,珠江口两岸的开发力度大,人类活动对珠江口生态系统的影响作用显著,出现了明显的富营养化现象。迄今,不少学者已对珠江口大、中型浮游动物的种类和数量进行了研究(李开枝等,2005;Li et al,2006;方宏达等,2009),但鲜见有关于中、小型浮游动物生态学研究的报道(Tan et al,2004)。本研究通过分析2009年的珠江口中、小型浮游动物种类、数量及其空间分布等生态学特征,有助于更好地揭示珠江口生态系统的结构和功能现状。
1 材料与方法
1.1 调查时间与站位
2009年的8月对珠江口海域18个站位进行了生物和理化因子的采样调查(图1)。
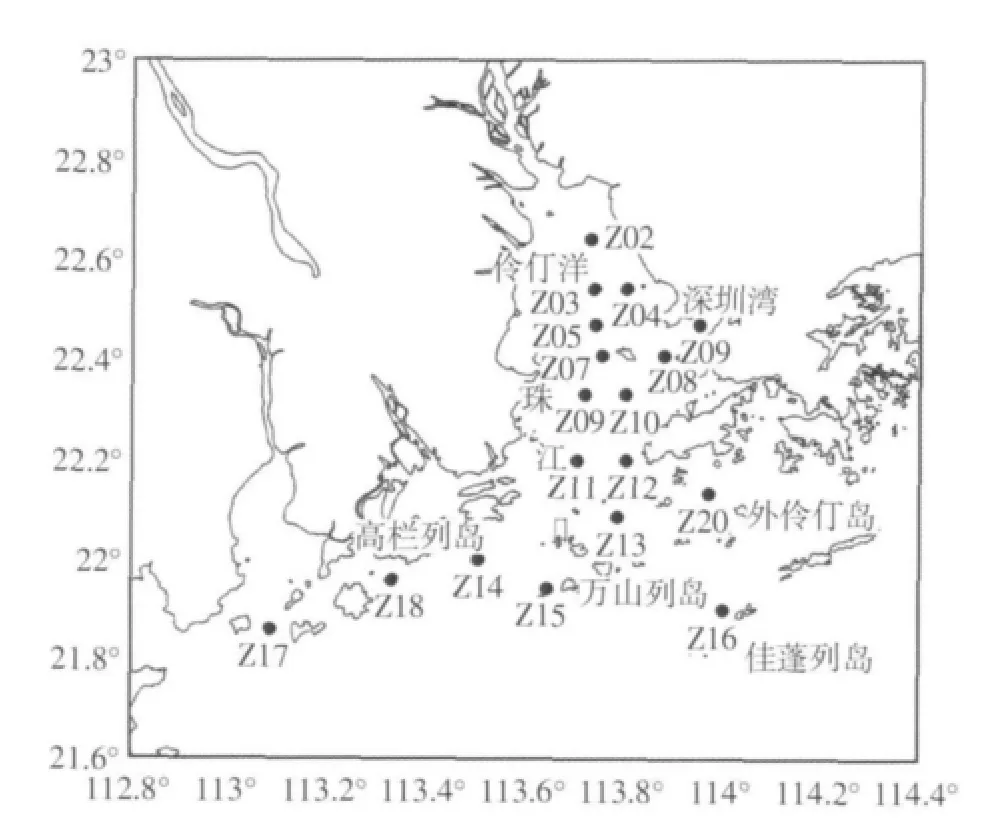
图1 采样站位图
1.2 现场采样与样品分析
浮游动物样品采用浅水Ⅱ型浮游生物网(网目孔径0.160 mm,网口内径31.6 cm,网长140 cm)由底层至表层进行垂直拖网一次。采集到的样品用5%的甲醛溶液固定,带回实验室分析鉴定。浮游动物采集、处理、计数和计算等采用《海洋调查规范第六部分:海洋生物调查》(GB 12763.6-2007,2007)提供的方法进行。主要环境因子(水温、盐度、叶绿素a浓度)数据引用其它专业同步测定结果。
1.3 数据处理
物种多样性指数采用Shannon-Weaver(Shannon,1948)指数(H′)(Shannon,1948)具体公式如下:
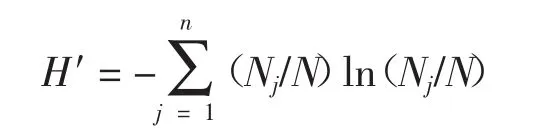
式中:n为群落中的物种数,N为群落中所有种的数量,Nj为群落中第j种的数量。
均匀度指数采用Pielou(1966)均匀度指数(J′)(Pielou,1966),计算公式如下:
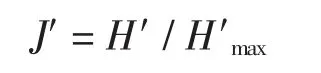
式中:H′为群落实测的物种多样性指数,H′max为群落的理论最大物种多样性指数log2S,S为群落中所有种的数量。
优势度(Y)采用以下计算公式:
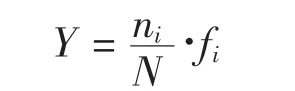
式中:ni为第i种的数量,fi为该种在各站出现的频率,N为群落中所有种的数量。当Y≥0.02时,该种为优势种(徐兆礼等,1989)。
调查海区各站位浮游动物种类数、个体数量、多样性指数、均匀度以及浮游动物及其优势种的个体数量与环境因子(水温、盐度和叶绿素a含量)的相关性采用Spearson相关系数(SPSS 12.0)进行分析。各站位之间浮游动物群落相似性分析,采用PRIMER 5.0软件包中的相似性聚类(Bray-Curtis Cluster)和多变量标序(Multidimensional Scaling,MDS)分析法,分析前先将数据通过log10(x+1)标准化(Li et al,2006)。
2 结果与讨论
2.1 种类组成
2009年8月对珠江口海域的调查中,共鉴定出终生浮游动物120种和阶段性浮游幼体5类。其中,桡足类最多,共58种,占总种数的48.3%;其次是水母类,共21种,占总种数的17.5%;被囊类10种,毛颚类8种,介形类4种,磷虾类5种,莹虾类3种,翼足类3种,端足类、栉水母类、枝角类、异足类和原生动物各2种。与我国其它近岸海区相比,珠江口中、小型浮游动物种类显著多于黄海南部,略丰富于珠江口的大、中型浮游动物种类(表1)。
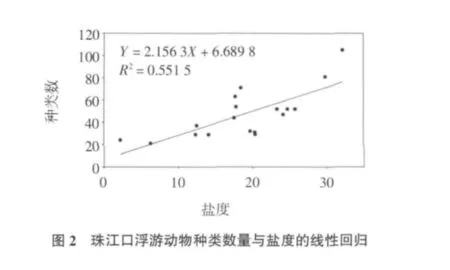
根据相关分析结果显示,调查海区浮游动物的种类数与水层平均盐度和温度呈显著相关关系,其中与盐度(R=0.743,P<0.01)呈显著正相关,与温度呈(R=-0.798,P<0.01)显著负相关。盐度是影响珠江口浮游动物的种类组成的重要环境因子,过去研究表明,珠江口大、中型浮游动物的种类数从内河口向外河口逐渐增加的趋势(李开枝等,2005;Li et al,2006;方宏达等,2009)。本研究显示,珠江口中、小型浮游动物的种类数也是随着盐度的增加而增加(图2),在河口顶部低盐区一般只出现一些的半咸水种,向外随盐度逐渐增加,出现河口种、近岸种和外海种共存,种类组成逐渐丰富。在表层盐度小于10的区域(Z02-Z07站),只出现了少量原生动物、水母类、桡足类、枝角类、异足类、莹虾类、磷虾类、毛颚类和被囊类,在盐度大于10的区域开始出现介形类、翼足类和端足类,到了盐度大于20的区域(Z10、Z11、Z14-Z16)水母类、桡足类、介形类、翼足类、端足类和毛颚类种类明显增多。调查期间,珠江口平均水温为28.97℃,其中表层平均为29.71℃,底层平均为28.23℃。夏季受温度较低的外海水从底层入侵的影响,低温区主要位于外海一侧的底层区域。水温与浮游动物种类数的负相关关系与沿岸流和外海水入侵带来多种生态类群的浮游动物种类有关。
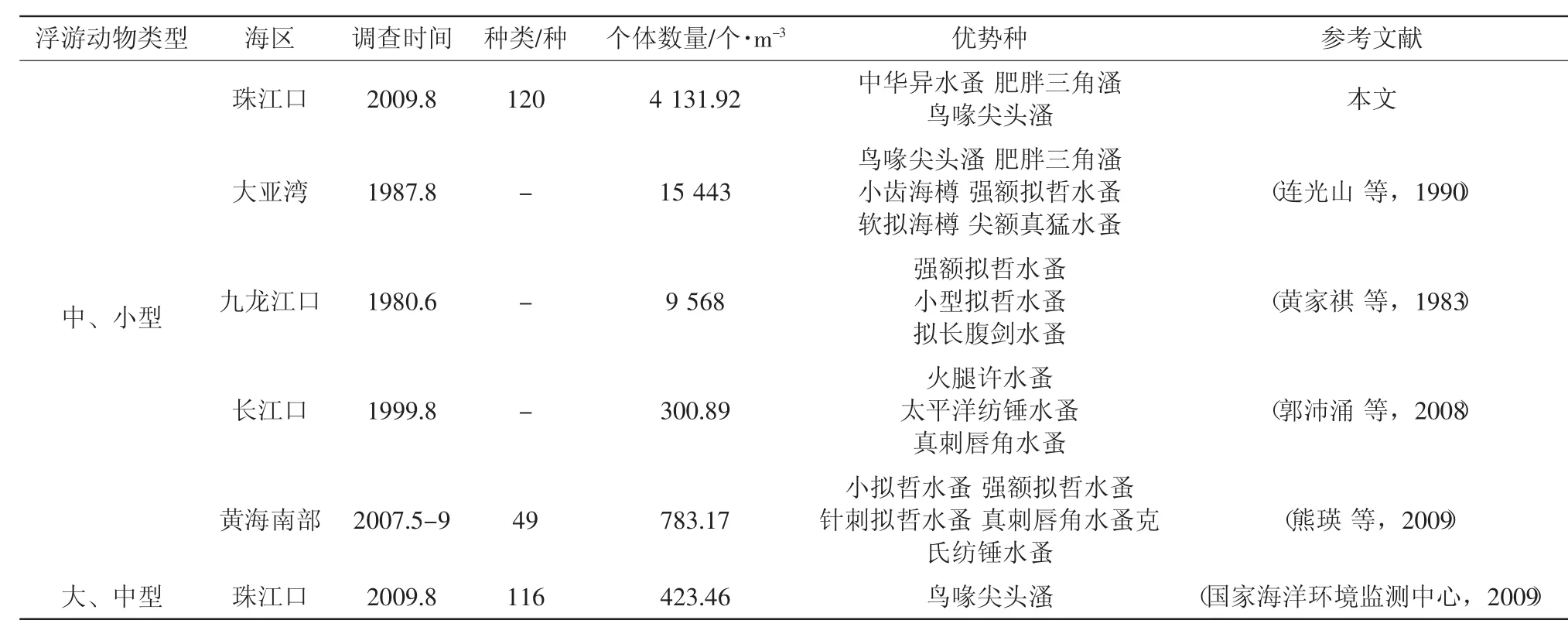
表1 珠江口与其它海区中、小型浮游动物种类数、数量和优势种及其与珠江口大、中型浮游动物的比较
2.2 生态类群
本研究中,珠江口浮游动物种类较多,依照其生态习性和地理分布主要分为以下4个生态类群(李开枝等,2005;Li et al,2006;方宏达 等,2009;郑重等,1984;张金标等,2003;李开枝等,2007)。
河口类群,主要生活在河口低盐水域,种类不多但数量较大。出现的代表种有中华异水蚤(Acartiella sinensis)、火腿许水蚤(Schmackeria poplesia)、球状许水蚤(Schmackeria forbesi)、刺尾纺锤水蚤(Acartia spinicauda)等。
近岸类群,主要生活在近岸盐度较低的水域。该类群种类较多、数量较大,是珠江口夏季的优势类群。其中,近岸广布类群的代表种为鸟喙尖头溞(Penilia avirosstris)、小拟哲水蚤(Paracalanus parvus);近岸暖水类群有肥胖三角溞(Evadna tergestina)、亨生莹虾(Lucifer hanseni)、半球美螅水母(Clytia hemisphaerica)、球型侧腕水母(Pleurobrachia globosa)、双生水母(Diphyes chamissonis)、弱 箭 虫(Sagitta delicata)和 尖 尾 海 萤(Cypridina acuminata)等。
外海类群,会随潮汐的涨落进入河口区,出现数量较少,但在靠近外海一侧种类较多。其中,外海暖水性类群的代表种有裂颏蛮虫戎(Lestrigonus schizogeneios)、角明螺(Oxygyrus keraudreni)、卵形光水蚤(Lucicutia ovalis)。外海广布类群代表种有普通波水蚤(Undinula vulgaris)、四叶小舌水母(Liriope tetraphylla)、半口壮丽水母(Aglaura hemistoma)、长吻纽鳃樽(Brooksia rostrata)等。
广温广盐类群,对温度和盐度的适应范围较广,代表种类为肥胖箭虫(Sagitta enflata)、小拟哲水蚤(Paracalanus parvus)、微驼隆哲水蚤(Acrocalanus gracilis)等。
2.3 优势种与群落结构
研究期间,珠江口海区浮游动物的优势种见表1。根据优势度计算结果,2009年夏季珠江口海区浮游动物的优势种有中华异水蚤、肥胖三角溞和鸟喙尖头溞,其优势度分别为0.18、0.11和0.06。从3种优势种的平面分布来看,中华异水蚤的平面分布与肥胖三角溞、鸟喙尖头溞的具有显著差异。中华异水蚤的数量从河口顶部向外呈下降的趋势,到了靠近外海的站位(Z14-Z18)则完全消失;而肥胖三角溞和鸟喙尖头溞的数量分布趋势则相反(图3)。优势种的空间变化表明珠江口浮游动物群落有较明显的空间异质性,其中由径流和外海水共同作用引起的盐度平面变化是关键影响因子之一。根据相关分析,中华异水蚤个体数量均与盐度呈显著负相关(R=-0.554,P<0.05),与该种为典型河口类群的生态习性有关。肥胖三角溞和鸟喙尖头溞是近岸类群的代表种,本研究中它们的个体数量与环境因子无显著相关关系,但二者个体数量却呈显著的正相关关系(R=0.996,P<0.01),表明肥胖三角溞和鸟喙尖头溞生态位较为接近。此外,浮游动物的生理生态特征是河口浮游动物群落结构变化的内在因素。如:鸟喙尖头溞可通过孤雌生殖使个体数量激增,当环境条件合适时能形成爆发性增殖,是珠江口夏季常见的优势种(Li et al,2006;方宏达等,2009)。
本研究未发现浮游动物优势种的分布与叶绿素a浓度有显著相关性,显示饵料浓度不是影响夏季珠江口浮游动物数量变动的主要原因,与大、中型浮游动物的相关研究结果相似(方宏达等,2009;Huang et al,2004;熊瑛 等,2012)。
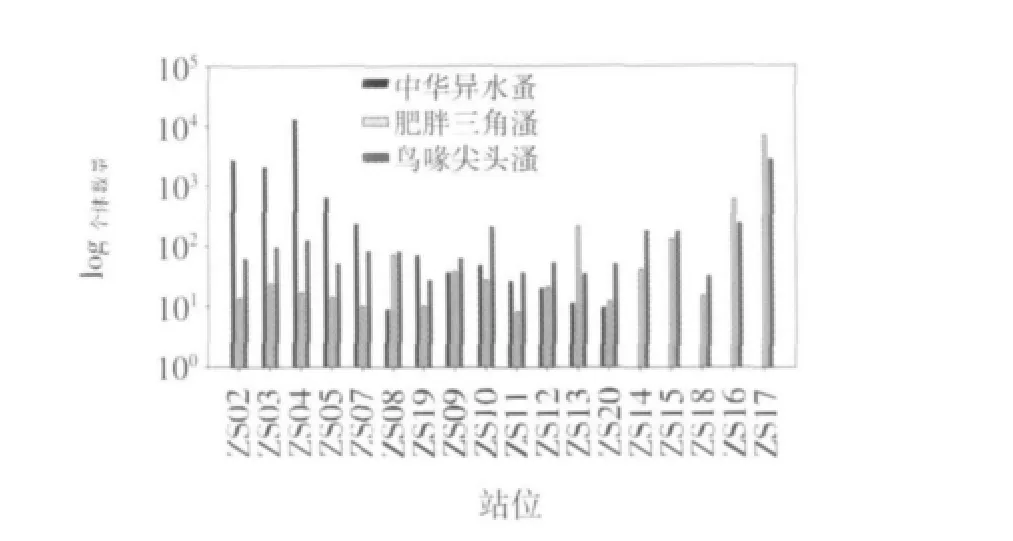
图3 珠江口浮游动物优势种在各站位个体数量(个/m3)的对数值
夏季珠江口处于丰水期,整个调查海域表层受珠江冲淡水主导,表层平均盐度只有14.4,此时浮游动物的优势种为河口种和近岸种。根据聚类分析结果显示,夏季珠江口浮游动物主要分为两大类:河口群落和近岸群落。河口群落集中分布在河口顶部表层盐度<5的区域(Z02、Z03、Z04),其它区域属于近岸群落(图4)。聚类分析结果主要与各类优势种的数量比例变动有关。在河口群落分布的区域,中华异水蚤的平均数量比例高达56%,该区域还出现大量典型河口种球状许水蚤(Schmackeria forbesi)和火腿许水蚤(Schmackeria poplesia)。在近岸群落分布区,肥胖三角溞和鸟喙尖头溞合共的平均数量比例为18%,而中华异水蚤在该群落中的数量比例下降至5%。其中,在Z15、Z16和Z17站,肥胖三角溞和鸟喙尖头溞大量繁殖,二者平均数量比例高达44%,因此在聚类分析中成为了近岸群落中一个较为特殊的分支(图4)。
2.4 多样性指数和均匀度
本研究中,珠江口浮游动物的多样性指数和均匀度的平均值分别为3.24和0.59,均略低于大、中型浮游动物的相应值(Li et al,2006;方宏达等,2009;Shannon,1948;Pielou,1966;徐兆礼等,1989;连光山等,1990;黄家祺等,1983;郭沛涌等,2008;国家海洋局南海环境监测中心,2009)。多样性指数的变化范围在1.18~4.48之间,均匀度的变化范围在0.27~0.87之间。多样性指数的高值区出现在靠近外海一侧的Z15站,此处盐度较高,多种生态类群的种类在此区域最为丰富(图2)。均匀度则在调查海区中部出现高值,该区优势种的数量优势较低(图2),不同种间的数量分布较为均匀。多样性指数的低值区主要出现在河口顶部,该区盐度低,浮游动物种类数较少(图2),以典型河口种为主,多样性指数小于2.00,加上中华异水蚤大量出现(图3),其他种类出现的数量较少,因此均匀度也在该区出现低值,小于0.5。其他区域多样性指数和均匀度分别大于2.50和0.50。此外,中华异水蚤的个体数量与多样性指数和均匀度呈显著负相关(R=-0.668,P<0.01;R=-0.604,P<0.01),表明中华异水蚤在局部区域的大量出现导致多样性指数和均匀度的下降。多样性指数和均匀度都与盐度呈显著正相关关系(R=0.773,P<0.01;R=0.593,P<0.01),与浮游动物种类数随盐度的增加和增加(图2)以及优势种的数量比例下降有关。
2.5 个体数量及其分布
珠江口浮游动物的个体数量变化较大,表现出明显的空间异质性。调查期间,浮游动物平均个体数量为4 131.92个/m3,与长江口和黄海南部相比要高得多,但不及大亚湾和九龙江口在上世纪80年代的水平(表1)。此外,与珠江口大、中型浮游动物的个体数量相比,中、小型浮游动物的个体数量也要高得多(表1)。本次调查,个体数量的最大值出现在河口顶部(图5),该区中华异水蚤大量出现。个体数量的最小值则出现在咸、淡水交汇处(Z11、Z14、Z15和Z18站,图1),该区水质层化现象明显,盐度垂直变化梯度大,叶绿素a含量也在该区出现低值,对浮游动物可能造成环境不稳定和食物不足的消极影响。这种分布状况与2005年夏季珠江口大、中型浮游动物的分布情况相似(方宏达等,2009)。另外,浮游动物个体数量在崖门口外的黄茅海水域(Z17站)出现高值,此处枝角类出现爆发性增殖,肥胖三角溞和鸟喙尖头溞数量均在该站达到高峰,占该站浮游动物总数的64%。该站叶绿素a含量(2.07mg/m3)明显高于临近的Z18站(0.18mg/m3)和Z14站(0.10mg/m3),为枝角类的繁殖提供了物质基础。
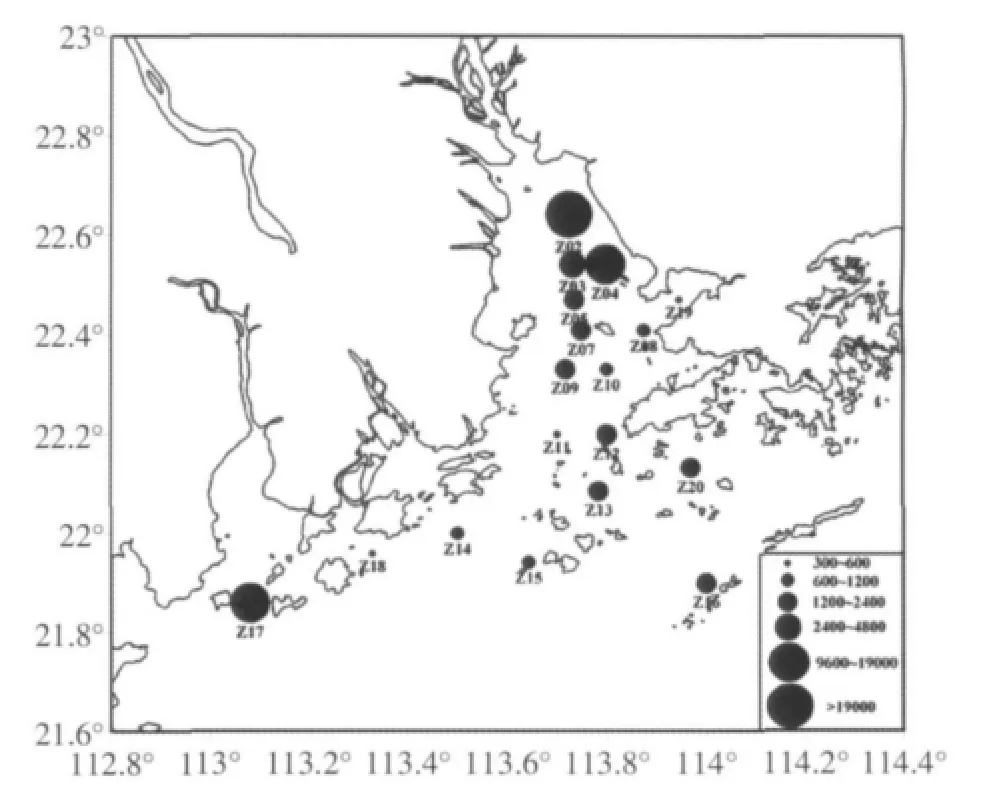
图5 珠江口浮游动物个体数量的空间分布(个/m3)
3 结论
(1)2009年8月调查共记录终生浮游动物120种和阶段性浮游幼体5类,其中桡足类种类最丰富,有58种,其次时水母类,有21种。海区优势种为中华异水蚤、肥胖三角溞和鸟喙尖头溞。聚类分析结果表明,珠江口顶部的浮游动物群落为河口群落,以中华异水蚤占绝对优势;其他区域为近岸群落,以肥胖三角溞和鸟喙尖头溞占优势。
(2)珠江口浮游动物平均个体数量4131.92个/m3,在河口顶部最高,咸淡水交汇区数量最低。黄茅海水域肥胖三角溞和鸟喙尖头溞出现爆发性增殖,可能与该处叶绿素含量较高有关。
(3)浮游动物物种多样性指数和均匀度的平均值分别为3.24和0.59,在靠近外海一侧和调查海域中部较高,河口顶部较低。其中,盐度与种类数呈显著正相关关系,而中华异水蚤的个体数量则和盐度呈显著负相关关系。可见,盐度和浮游动物本身的生态特征是影响浮游动物群落结构的主要因素。
Huang L M,Jian W J,Song X Y,et al,2004.Species diversity and distribution for phytoplankton of the Pearl River estuary during rainy and dry seasons.Marine Pollution Bulletin,49(7-8):588-596.
Li K Z,Yin J Q,Huang L M,et al,2006.Spatial and temporal variations of mesozooplankton in the Pear River estuary,China.Estuary,Coastal and Shelf Science,67:543-552.
Morales C E,Bedo H A,Harris P,et al,1991.Grazing of copepod assemblages in the north-east Atlantic:The importance of the small size fraction.Journal of Plankton Research,13:455-472.
Pielou E C,1966.The measure of diversity in different types of biological collections.Journal of Theoretical Biology,13:131-144.
Shannon C E,1948.A mathematical theory of communication.Bell System Technology Journal,27:379-423.
Tan Y H,Huang L M,Chen Q C,et al,2004.Seasonal variation in zooplankton composition and grazing impact on phytoplankton standing stock in the Pearl River Estuary,China.Continental Shelf Research,24(16):1949-1968.
GB12763.6-2007,2007,海洋调查规范第六部分:海洋生物调查.北京:中国标准出版社.
方宏达,朱艾嘉,董燕红,等,2009.2005~2006年珠江口浮游动物群落变化研究.台湾海峡,28(1):30-37.
郭沛涌,沈焕庭,刘阿成,等,2008.长江河口中小型浮游动物数量分布、变动及主要影响因素.生态学报,28(8):3517-3526.
国家海洋局南海环境监测中心,2009,2009年珠江口生态监控区监测报告.广州:国家海洋局南海环境监测中心.
黄加祺,1983.九龙江口大、中型浮游动物的种类组成和分布,厦门大学学报(自然科学版),22,1:88-95.
李开枝,尹健强,黄良民,等,2005.珠江口浮游动物的群落动态及数量变化.热带海洋学报,24(5):60-68.
李开枝,尹健强,黄良民,2007.河口浮游动物生态学研究进展.海洋科学,31(3):72-75.
连光山,蔡秉及,林玉辉,等,1990.大亚湾浮游动物生物量和密度的分布,大亚湾生态文集(II),海洋出版社,北京.
王荣,张鸿雁,王克,等,2002.小型桡足类在海洋生态系统中的功能作用.海洋与湖沼,33(5):453-460.
熊瑛,王云龙,刘培廷,等,2012.辐射沙脊群南部浮游动物的生态特征.海洋通报,31(2):181-188.
熊瑛,王云龙,汤建华,等,2009.黄海南部大型水母暴发区中小型浮游动物生态特征.生态学杂志,28(10):2063-2068.
徐兆礼,陈亚瞿,1989.东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系.生态学杂志,8(4):13-15.
张金标,黄将修,连光山,2003.台湾南湾区秋末冬初海樽类的种类组成和数量分布.海洋通报,22(6):9-16.
郑重,李少菁,许振祖,1984.海洋浮游生物学.北京:海洋出版社.
