灵石山米槠林优势树种不同年龄叶片养分特征
2012-07-30王英姿
王英姿
(福州市林业局,福建 福州350005)
因为对养分的敏感性以及易于采集等特点,所以叶片功能性状的研究成为学者关注的焦点[1]。植物的养分特征与其生长能力和生产力密切相关[2]。植物叶片养分含量特征研究对于森林树种养分状况评估、森林退化地区合适的乡土树种的选择以及森林生态系统可持续经营具有重要意义[3-5]。目前,更多的研究是关注植物养分过程而不是树种养分特征状况[6]。有关中国亚热带阔叶林叶片养分特征的研究仅见于林植芳等[7]、莫江明等[8]对于鼎湖山常绿阔叶林的研究报道,表明不同生活型植物对矿质元素的吸收和积累效应不同;磷和镁可能是限制南亚热带常绿阔叶林植物生产力的最重要的营养元素。有关热带雨林叶片养分含量的研究表明,物种多样性和环境异质性使得叶片养分含量变化范围变大[9-10]。Vitousek等[3]研究表明:植物组织的养分含量很可能反映了土壤肥力的影响,生长在不同土壤肥力中的同一树种叶片养分含量通常是相似的。Masunaga等[10]对印度尼西亚苏门答腊岛植物养分特征的研究提供了有关养分特征研究的新视角,他们将某一优势种养分对数与该组优势种养分对数的平均值和标准差作比较,将植物种分为该养分的“积累种”(accumulator)和“排斥种”(excluder),并得出结论,植物种养分特征的高变异性反映了研究地植物种较高的多样性。米槠Castanopsis carlesii林是中国东部湿润亚热带地区常绿阔叶林的重要建群种,也是世界上罕见的植被类型[11-12]。米槠适应性强,分布广,培肥土壤、涵养水源的能力较强,米槠群落结构较为复杂,物种相对丰富,近年来笔者所在课题组首次对米槠林优势种叶片养分结构、叶绿素等叶属性特征沿海拔的变化进行研究并报道[13-15],表明叶属性是植物长期适应环境的结果,但有关优势种叶片养分特征的探讨还不够深入。本研究进一步对灵石山不同海拔米槠林优势种不同年龄叶片的养分特征进行研究,旨在分析灵石山米槠林不同优势种的环境适应策略及对环境的适应能力,为选择乡土阔叶树种造林提供理论依据。
1 研究区概况
灵石山国家森林公园位于闽中福清市西部,25°40′N,119°13′E,总面积为2 275.0 hm2,其中有林地面积为2 040.5 hm2,森林覆被率93.5%,属南亚热带气候带闽东南沿海海洋性季风气候区域。全年平均气温为19.7℃,7月平均气温29.0℃,1月平均气温7.0℃,极端最高气温34.0℃,极端最低气温0.0℃。年均日照时数为2 000 h,无霜降期340~360 d,有雾日20~30 d。年平均降水量为1 780 mm,雨季多在3-6月,相对湿度86%。森林植被保持良好,公园内水质甚佳,清澈透明,各山涧小溪水源常年不断。土壤大多为花岗岩发育的酸性土壤,地带性土壤为红壤,其中海拔600 m以上为黄红壤。公园内土壤表层物理性能良好,土层深厚,腐殖质层较厚。在《福建植被区划》上,灵石山森林公园属于闽粤沿海丘陵平原南亚热带雨林区闽江口、鹫峰山南湿暖南亚热带雨林小区。海拔250 m以下为地带性植被,随着海拔的增高,森林植被依次为亚热带常绿阔叶林、山地矮林,间有马尾松Pinus massoniana林和竹林。
2 研究方法
2.1 调查与实验方法
根据灵石山国家森林公园米槠林的主要分布范围,结合全球定位系统(GPS)定位技术,采用样线和样地相结合的方法,于2008年3月在灵石山设9个不同海拔梯度,按海拔从低到高分别记为A1(157 m),A2(200 m),A3(242 m),A4(332 m),A5(442 m),A6(531 m),A7(632~662 m),A8(762 m),A9(842 m)。每一海拔高度视野外情况而定,选取1~2个20 m×20 m的典型样地,将各样地分成16个5 m×5 m的小样方,对各小样方内胸径≥2.5 cm的树种进行每木调查,实测胸径、树高、冠幅等指标,其中,树高用测高仪测定,冠幅为树冠最大直径和最小直径的平均值,直径用皮尺测量[16]。通过计算重要值确定优势种,A1~A9的所有优势种有米槠Castanopsis carlesii,闽桂润楠Machilus minkweiensis,鹅掌柴Schefflera octophylla,木荷Schima superba,微毛柃Eurya hebeclados,尖叶柃Eurya acuminatissima,罗浮栲 Castanopsis fabri,山杜鹃Rhododendron simisii,九节木Psychotria rubra,罗伞树Ardisia quin-quegona,围涎树Pithecellobium clypearia,赤楠Syzygium buxifolium,青冈栎Cyclobalanopsis glauca,密花树 Rapanea neriifolia,乌药Lindera aggregata,油茶Camellia oleifera,石栎Lithocarpus glaber,柏拉木Blastus cochinchinensis,肉实树Sarcosperma laurinum,黑壳楠Lindera megaphylla,厚叶冬青Ilex elmerrilliana,狗骨柴Diplospora dubia,八瓣糙果茶Camellia octopetala(表1)。然后,每样地中选择各优势树种接近平均胸径的标准木3株,在树冠的中上层分东西南北4个方向采集树枝,按叶鳞痕等生长状况将叶片分为1年生和2年生叶片分别500 g左右,封装后立刻带回实验室处理,并进行各种养分质量分数的测定[17-19]。

表1 灵石山米槠林不同海拔优势种Table 1 List of dominant species in Castanopsis carlesii forest at different altitude gradients
2.2 数据分析
实验所得数据采用SPSS 13.0软件中的Data EXPLOR进行对数正态分布检验,使用Excel绘制对数分布图及计算平均值和标准差。重要值=相对多度+相对频度+相对显著度[20]。
3 结果与分析
3.1 不同年龄叶片养分特征及其分布特征
由表2可以看出:米槠林优势种养分种间差异比较大,1年生叶片最低和最高的比值分别为:1∶17.9(氮),1 ∶6.7(磷),1 ∶15.0(钾),1 ∶11.2(钠),1 ∶21.1(钙),1 ∶4.3(镁); 1 ∶13.1(铁); 1 ∶5.1(锰); 1 ∶11.0(锌); 2年生叶片最低和最高含量的比值分别为:1 ∶7.0(氮),1 ∶6.0(磷),1∶12.8(钾),1 ∶14.4(钠),1 ∶13.3(钙); 1 ∶4.3(镁); 1 ∶7.29(铁); 1 ∶7.4(锰); 1 ∶15.1(锌),1 年生叶片养分以钙的种间差异最大,其次是氮,最小为镁;2年生叶片养分以锌的种间差异最大,其次为钠,最小为镁;不同年龄叶片养分差异最大的是氮、铁和钙,反映了植物种对养分元素的选择性吸收。
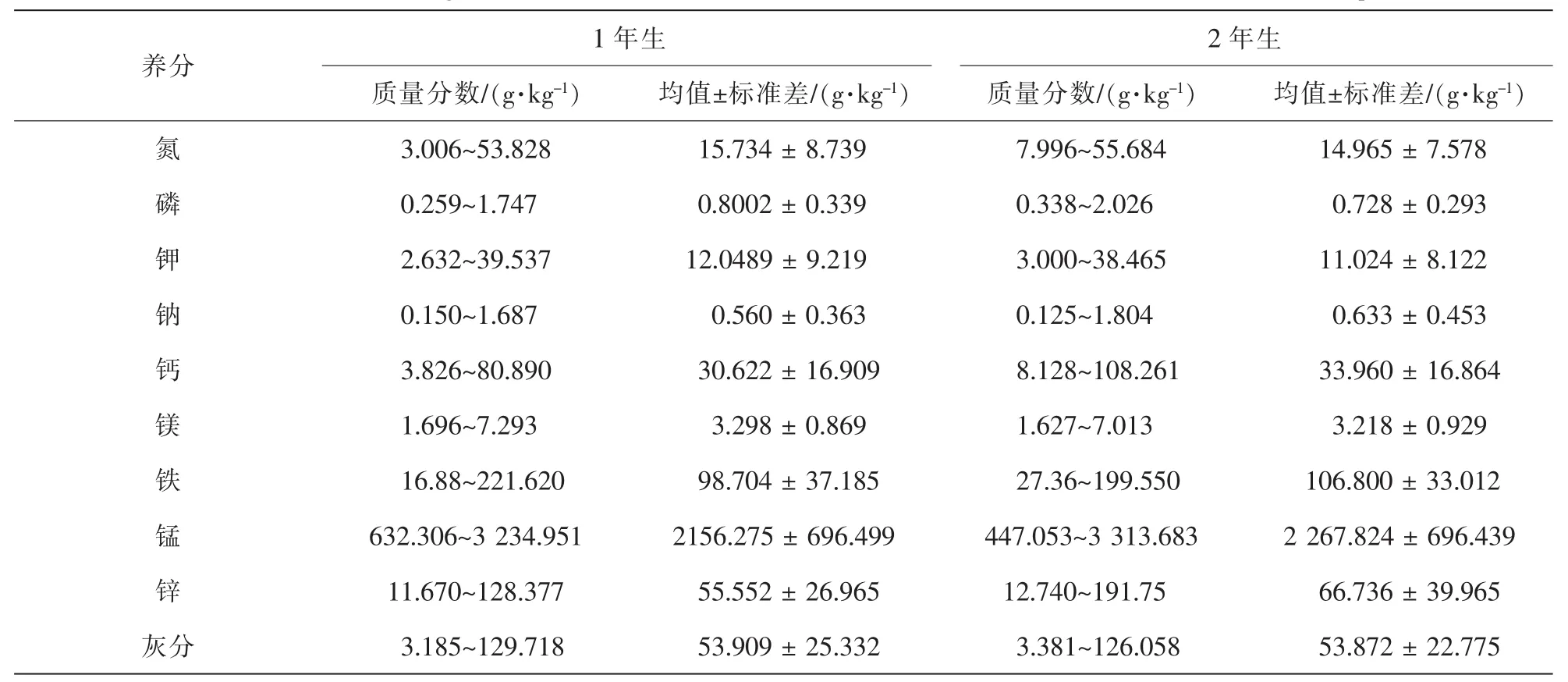
表2 灵石山米槠林优势树种不同年龄叶片养分特征Table 2 Concentration ranges and mean values of the of mineral nutrients in the leaves of the dominant tree species
对米槠林优势种不同年龄叶片的养分的频数分布进行分析,表明不符合正态分布,对各养分取以10为底的对数,然后进行正态分布Kolmogorov-Smirnov检验(表3),结果表明:不同年龄叶片磷、2年生叶片钠、2年生叶片钙、1年生叶片镁、1年生叶片铁、不同年龄叶片锌符合对数正态分布(图1)。氮和钾的对数频数分布图峰值多于1个,氮约在1.0,1.1,1.2,钾约在0.7,0.8,0.9,1.3,可以被解释为优势树种有不同的分组[4,19]。
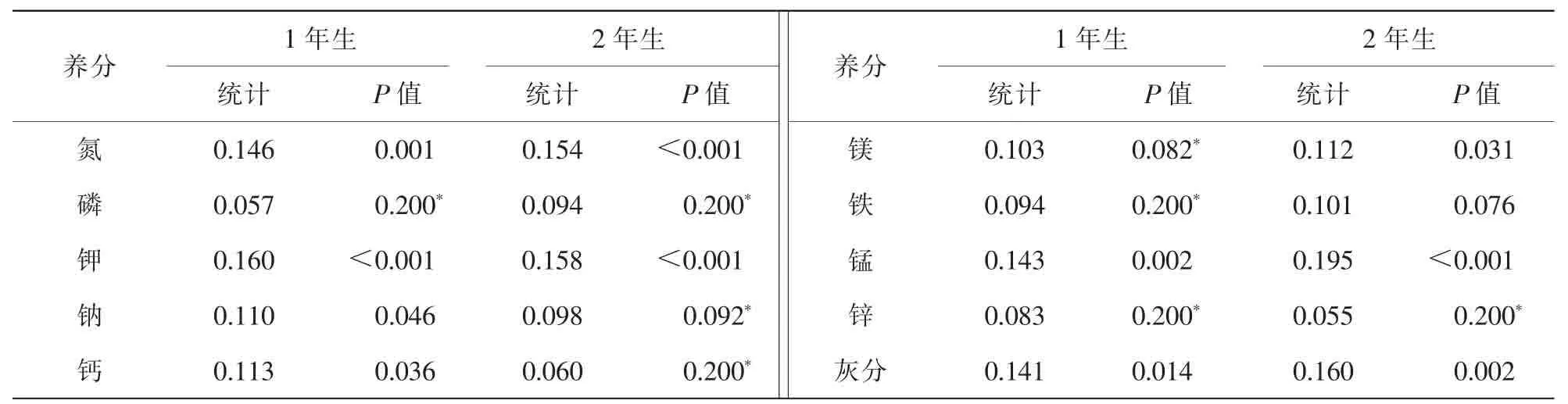
表3 灵石山米槠林不同年龄叶片养分对数正态分布检验Table 3 Test of logarithmic normal distribution of concentrations of mineral nutrients in the leaves of dominant species
3.2 灵石山米槠林优势种养分的分类
由叶片养分的频数分布图(图1)可以看出:叶片的养分范围可以分为3类:低、中、高,其中叶片养分居于中等范围内的优势种居多。图2~3为灵石山米槠林22种优势种各种养分特征,将优势种的养分分为4类。依据为:养分的对数①小于(X-2SD),②介于(X-2SD)和X之间,③介于X和(X+2SD)之间,④大于(X+2SD)。X和SD分别为养分对数的平均值和标准差。对于某一优势种,其养分对数小于(X-2SD)被称为该种养分的“排斥种 ”(excluder),而养分的对数大于(X+2SD)被称为该种养分“累积种”(accumulator)[4]。
图2~3可以直观地看出22种优势种各种养分特征。厚叶冬青1年生叶片磷较低,对磷有“排斥”效应,青冈栎2年生叶片锌较低,显示出对锌的“排斥”效应。不同年龄鹅掌柴叶片钾比较高,被分为钾“累积”种,2年生狗骨柴叶片对钠、1年生狗骨柴叶片对钾有明显的“累积”效应,不同年龄围涎树叶片氮、2年生石栎叶片氮明显高于其他优势种,有较强的氮“累积”效应。其他优势种叶片养分均为第2类或第3类,但是不同年龄叶片对同种养分的“累积”效应或者说其在同龄叶片中的分类等级不尽相同。总体来说,优势种2年生叶片养分质量分数对数大于平均值即处于第3分类等级的比例大于1年生叶片。
石砾、围涎树不同年龄叶片的氮均比较高。除氮和钠以外,九节木不同年龄叶片各种养分均比较高。鹅掌柴不同年龄叶片磷、钾、钠、镁、锌较其他优势种高。微毛柃、细齿柃和山杜鹃叶片钙比较高。微毛柃、细齿柃和八瓣糙果茶叶片铁以及锰质量分数均较高。相对于其他优势种,密花树叶氮较低;厚叶冬青叶磷比较低,特别是1年生叶片磷在所有优势种中最低;青冈栎叶片锌较低,特别是2年生叶片,居于第4类为锌的排斥种。
4 小结与讨论
植物的养分特征受营养吸收能力、土壤养分的有效性以及其他环境因子的影响[5,10,21],它不仅能揭示植物种的特性,同时还能反映植物与环境之间的关系。
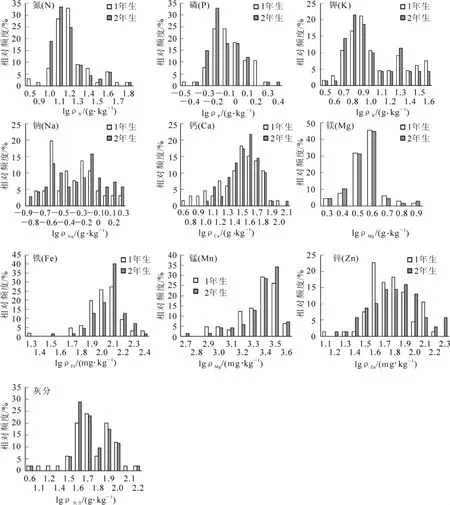
图1 灵石山米槠林不同年龄叶片养分含量对数分布频数图Figure 1 Frequency distribution of the logarithmic concentrations of mineral nutrients in the leaves of dominant species in Castanopsis carlesii forest
灵石山米槠林优势种不同年龄叶片磷、2年生叶片钠和钙、1年生叶片镁和铁、不同年龄叶片锌均符合对数正态分布。钙和氮分布范围较宽,优势种间差异性较高,磷和镁分布范围比较窄,优势种间差异性较低。与陈灵芝等[22]对中国各气候带代表植物叶片养分总结的结果相比,本研究中,氮、钾和镁与中国各气候带的平均水平相似,钙则显著高于各气候带的平均水平,磷则显著低于各气候带的平均水平,占各气候带总平均水平的54%。叶片是植物进行光合作用的重要场所,它的营养元素特征最具有代表性并最能反映群落的生境条件,磷是很多热带森林土壤中的限制性养分[23-24],被认为是热带雨林生产力的限制性因子[25]。莫江明等[8]研究也表明:磷是鼎湖山南亚热带常绿阔叶林植物生产力的限制因素。通过本研究结果可以推断磷是灵石山米槠林植物生长的限制性因子。有关森林叶片钠的研究很少,钠在农作物上的作用研究表明,钠作为功能元素具有部分替代钾营养的作用[26-27]。本研究中,优势种2年生叶片钾平均小于1年生叶片,而2年生叶片钠平均大于1年生叶片,说明钠在优势种生长过程中钠起了促进作用。

图2 基于养分质量分数分级的灵石山米槠林不同优势种1年生叶片养分特征Figure 2 Nutrient characteristics in one-year-old leaves of the dominant tree species based on four concentration classes or grade
基于养分特征的优势种养分分类方法,可以较明显地表明米槠林优势种各养分的差异性。优势种米槠各种养分居于第2类即小于平均值的养分的比重比较大,这是物种对环境的生存适应策略[28]。树种对于某种养分的吸收累积能力受遗传及环境因素共同影响[4,29-30],相对于叶片的解剖结构,植物根系的养分选择吸收系统对于养分元素的影响更大[31]。本研究中,围涎树和石栎有较高的氮吸收能力,而鹅掌柴对于除钙、铁以外的其他养分的吸收能力都较高,九节木对于各种养分的吸收能力都比较高。微毛柃、细齿柃对微量元素铁、锰、锌的吸收能力均较高。厚叶冬青、青冈栎分别较低的磷、锌表明它们可以耐受这种养分缺乏的生境,可能是贫瘠环境下造林的适合树种。优势种不同的养分吸收机制值得进一步研究。
[1] van den DRIESSCHE R.Prediction of mineral nutrient status of trees by foliar analysis [J].Bot Rev,1974,40:347-395.
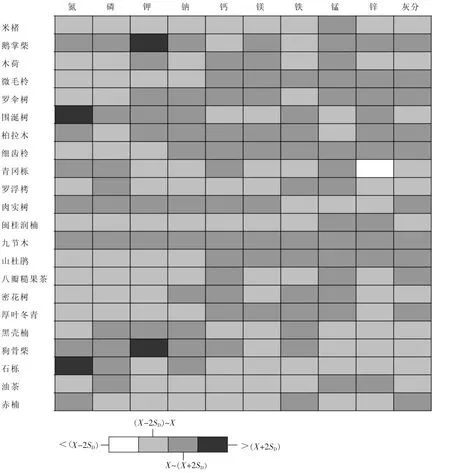
图3 基于养分质量分数分级的灵石山米槠林不同优势种2年生叶片养分特征Figure 3 Nutrient characteristics in two-year-old leaves of the dominant tree species based on four concentration classes or grade
[2] GREGOIRE N,FISHER R.Nutritional diagnoses in loblolly pine(Pinus taeda L.) established stands using three different approaches [J].For Ecol Manage,2004,203(1/3):195-208.
[3] VITOUSEK P M,SANFORD R L.Nutrient cycling in moist tropical forests [J].Ann Rev Ecol Syst,1986,17:137-167.
[4] WU C C,TSUI C C,HSEIH C F,et al.Mineral nutrient status of tree species in relation to environmental factors in the subtropical rain forest of Taiwan [J].For Ecol Manage,2007,239(1/3):81-91.
[5] NUSSBAUM R,ANDERSON J,SPENCER T.Factors limiting the growth of indigenous tree seedlings planted on degraded rain forest soils in Sabah,Malaysia [J].For Ecol Manage,1995,74(1/3):149-159.
[6] ROTHE A,EWALD J,E.HIBBS D.Do admixed broadleaves improve foliar nutrient status of conifer tree crops? [J].For Ecol Manage,2003,172(2/3):327-338.
[7]林植芳,李双顺,孙谷筹,等.鼎湖山南亚热带地区植物的叶片矿质元素[J].生态学报,1989,9(4):320-323.LIN Zhifang,LI Shuangshun,SUN Guchou,et al.The mineral elements in leaves of plants of in south subtropical area of Mt.Dinghushan [J].Acta Ecol Sin,1989,9(4):320-323.
[8]莫江明,张德强,黄忠良,等.鼎湖山南亚热带常绿阔叶林植物营养元素含量分配格局研究 [J].热带亚热带植物学报,2000,8(3):198-206.MO Jiangming,ZHANG Deqiang,HUANG Zhongliang,et al.Distribution pattern of nutrient elements in plants of Dinghushan lower subtropical evergreen broad-leaved forest[J].J Trop Subtrop Bot,2000,8(3):198-206.
[9] MASUNAGA T,KUBOTA D,HOTTA M,et al.Nutritional characteristics of mineral elements in leaves of tree species in tropical rain forest,west Sumatra,Indonesia [J].Soil Sci Plant Nutr,1998,44:315-329.
[10] MASUNAGA T,KUBOTA D,WILLIAM U.Spatial distribution pattern of trees in relation to soil edaphic status in tropical rain forest in west Sumatra,Indonesia(I) distribution of accumulation [J].Tropics,1998,7:209-222.
[11]闫淑君,洪伟,吴承祯,等.武夷山米槠林优势种群的竞争格局及动态模拟[J].福建林学院学报,2002,22(1):1-3.YAN Shujun,HONG Wei,WU Chengzhen,et al.Competitive pattern and dynamics simulation of dominant population in Castanopsis carlessi forest in Wuyishan Mountains [J].J Fujian Coll For,2002,22(1):1-3.
[12]闫淑君,洪伟,吴承祯,等.武夷山天然米槠林优势种群结构与分布格局[J].热带与亚热带植物学报,2002,10(1):15-21.YAN Shujun,HONG Wei,WU Chengzhen,et al.The structure and distribution pattern of dominant populations in Castanopsis carlesii community [J].J Trop Subtrop Bot,2002,10(1):15-21.
[13]王英姿,洪伟,吴承祯,等.灵石山米槠林优势种群不同叶龄叶属性的研究[J].福建林学院学报,2009,29(3):203-209.WANG Yingzi,HONG Wei,WU Chengzhen,et al.Research on leaf traits for different age leaves of dominant species in Castanopsis carlesii forests in Lingshishan National Forest Park [J].J Fujian Colle For,2009,29(3):203-209.
[14] WANG Yingzi,HONG Wei,WU Chengzhen,et al.Research on the variation of uneven-aged leaf SPAD values and the correlation between SPAD and chlorophyll,nitrogen concentration of dominant species in Castanopsis carlesii forsests in Lingshihan National Forest Park [J].J For Res,2009,29(3):203-209.
[15]王英姿,洪伟,吴承祯,等.灵石山米槠林优势种群不同年龄叶片叶绿体色素沿海拔梯度的变化分析[J].林业科学,2010,46(11):43-51.WANG Yingzi,HONG Wei,WU Chengzhen,et al.Variation of chloroplast pigments of various age leaves along elevation gradients of dominant species in Castanopsis carlesii forests in Lingshishan National Forest Park [J].Sci Silv Sin,2010,46(11):43-51.
[16]罗玲,廖超英.榆林沙区樟子松冠幅与胸径的相关关系分析[J].安徽农学通报,2007,13(24):92-97.LUO Ling,LIAO Chaoying.Research on the correlation between crown width and DBH of Pinus sylvestris var.mongolica in Yulin desert area [J].Anhui Agric Sci Bull,2007,13(24):92-97.
[17]祁建,马克明,张育新.北京东灵山不同坡位辽东栎Quercus liaotungensis叶属性的比较[J].生态学报,2008,28(1):122-128.QI Jian,MA Keming,ZHANG Yuxin.Comparisons on leaf traits of Quercus liaotungensis Koidz.on different slope positions in Dongling Moutain of Beijing [J].Acta Ecol Sin,2008,28(1):122-128.
[18]李永华,罗天祥,卢琦,等.青海省沙珠玉治沙站17种主要植物叶性因子的比较[J].生态学报,2005,25(5):994-999.LI Yonghua,LUO Tianxiang,LU Qi,et al.Comparisons of leaf traits among 17 major plant species in Shazhuyu Sand Control Experimental Station of Qinghai Province [J].Acta Ecol Sin,2005,25(1):994-999.
[19]国家林业局.LY/T 1210-1275-1999中华人民共和国林业行业标准:森林土壤分析方法[S].北京:中国标准出版社,1999:274-298.
[20] MASUNAGA T,KUBOTA D,HOTTA M,et al.Distribution characteristics of mineral elements in trees of tropical rain forest,West Sumata,Indonesia [G]//SCHULTE A,RUHIYAT D.Soil of Tropical Forest Ecosystems.Heidel-berg:Springer,1998:168-174.
[21]王伯荪.植物群落学[M].北京:高等教育出版社,1987:42.
[22]陈灵芝,黄建辉,严昌荣.中国森林生态系统养分循环[M].北京:气象出版社,1997:214.
[23] SUSAYA J P,ASIO V B.Status of phosphorus in the rain forest of Mt.Pangasugan,Leyte,Philippines [J].Ann Trop Res,2005,27:70-82.
[24] ZIKELI S,ASIO V B,JAHN R.Nutrient characteristics of soil in the rain Forest of Mt.Pangasugan,Leyte,Philippines [J].Ann Trop Res,2000,22:78-88.
[25] WARDLE D A,WALKER L R,BARDGETT R D.Ecosystem properties and forest decline in contrasting long-term chronosequences [J].Science,2004,305:509-513.
[26]吴延寿,尹建华,彭志勤,等.钠钾交互作用下水稻生长和营养元素吸收特征研究[J].安徽农业科学,2009,37(10):4456-4458.WU Yanshou,YIN Jianhua,PENG Zhiqin,et al.Study on rice growth and characteristics of nutrient elements absorption under interaction of K and Na [J].J Anhui Agric Sci,2009,37(10):4456-4458.
[27]刘国栋,刘更另.水稻不同基因型中Ca,Na对K的部分替代作用[J].作物学报,1996,22(3):313-320.LIU Guodong,LIU Gengling.Partial substitution function of Ca and Na for K in different genotypic rice [J].Acta A-gron Sin,1996,22(3):313-320.
[28] WRIGHT I J,REICH P B,WESTOBY M,et al.The world wide leaf e-comics spectrum [J].Nature,2004,42(8):821-827.
[29] DÍAZ S,CABIDO M.Vive la difference:plant functional diversity matters to ecosysten processes [J].Trends Ecol Evol,2001,16(11):646-655.
[30] MARTINS P,SAMPDRO L,MOREIRA X,et al.Nutritional status and genetic variation in the response to nutrient availability in Pinus pinaster:a multisite field study in Northwest Spain [J].For Ecol Manage,2009,258(7):1429-1436.
[31] GRUBB P J.Control of forest growth and distribution on wet tropical mountains:with special reference tomineral nutrition [J].Ann Rev Ecol Syst,1977,8:83-107.
