Effects of Phosphate Fertilizer on Cold Tolerance and Its Related Physiological Parameters in Rice Under Low Temperature Stress
2012-07-02HouLigangChenWenfuZhaoGuochenMaWeiQiChunyanLiuLiangandSunHongjiao
Hou Li-gang , Chen Wen-fu, Zhao Guo-chen, Ma Wei, Qi Chun-yan, Liu Liang, and Sun Hong-jiao
1 Rice Research Institute, Shenyang Agricultural University, Shenyang 100866, China;
2 Rice Research Institute, Jilin Academy of Agricultural Sciences, Gongzhuling 136100, Jilin, China
Introduction
Located in the East Asian monsoon region, China is one of the climate vulnerable areas in the world.According to statistics, only in the agricultural field,more than 4 800 million hectares of agricultural production were hit by meteorological disasters each year, which caused serious economic losses in grain production. Cold damage, as a current major agrometeorological disaster, led to China's annual losses of up to 50 to 10 billion kg only in rice production (Wang,2008). Analysis on the meteorological data of the last 50 years (1951 to 2000)in Jilin Province shows that there is an annual average rise in temperature, but in the main rice growing season, regional and stage cold damage phenomena still occur from time to time, and tend to be frequent and severe, and thus result in a year of cold damage (Yu et al., 2003). Phosphorus, one of the necessary nutritional elements for plant growth and development, usually participates in the activities of plant life in multiple ways. The growing number of studies have shown that phosphorus can signif i cantly improve the resistance of the plant, especially in cold tolerance (Wang et al., 1999; Li et al., 2004; Wei et al.,2005; Chen et al., 2007). Years of practice has confirmed that low temperature can affect the rice phosphate absorption and inhibit the growth; while increasing P fertilizer can promote the growth of rice root and help to turn the seedlings green after transplanting, thereby improving rice yield. In recent years,studies on sugar cane (Wei et al., 2003), watermelon(Chen et al., 2007), winter wheat (Li et al., 2004)and tobacco (Wang et al., 1999)plants show that an appropriate increase in P fertilizer plays an important role in improving plant tolerance to cold. However,few studies are conducted on the physiological mechanism of how the increase of P fertilizer can enhance low temperature resistance of rice seedlings.Taking the mechanism as object, this study employed two cold tolerance rice varieties as materials to research the effects of increased P fertilizer on the rice seedlings under low temperature cold tolerance and related physiological characteristics. It aimed to further elucidate the physiological mechanism and provided a theoretical basis for phosphorus fertilizer-centered anti-cold-technical measures.
Materials and Methods
Tested materials
The test was conducted in the Breeding Base of Jilin Academy of Agricultural Sciences in 2010, taking different cold tolerance varieties Jijing 81 (chillingtolerance, Jijing 81)and Changbai 9 (chilling-sensitive, Changbai 9)as test materials.
Experimental design
Firstly, selected the seeds of the same degree in maturity and plumpness and then soaked them for potential bud in mid-April 2010. Secondly, selected seeds with similar bud growth and sowed them evenly in seedling trays which were mixed with different phosphates, with the sowing rate of 200 g · m-2per disk. Five phosphate fertilizer application rates (P2O5)-0, 25, 50, 75 and 100 g · m-2were divided into two groups, one for low temperature processing, another group was under normal temperature control, each group was repeated twice. Finally, irrigated enough water and covered them in the seedling greenhouse with reference to the high yield of rice cultivation practices in Jilin Province.
Temperature treatment
When the rice seedlings grew up to three leaves,removed the group under low temperature treatment to the artif i cial climate chamber (LT/ACR-2002, artif i cial climate chamber control system of Yishengtaihe Company)for continual low temperature stress for 5 days and the average daily temperature was 8℃;referred to the specif i c parameter settings in Table 1.The other group stayed outdoor whose temperature data was collected via IRRI weather recorder; the concrete temperature is shown in Fig. 1 (Gongzhuling,2010). During the processing, the average daily temperature was over 15℃ and water was given quantitatively.
Measurement items and methods
Evaluation of cold tolerance ability of rice seedlings
With reference to the approaches by Han et al. (2004)and Li et al. (2006)and combined with the actual situation of Jilin Province, the study identified and rated the ability of rice seedlings by evaluating how dry and red of the leaves. The criteria were as the followings: the first grade-the rate of dry and red leaves≤20%; the second grade-the rate of dry and red leaves was between 21%-30%; the third gradethe rate of dry and red leaves was between 31%-40%;the fourth grade-the rate of dry and red leaves was between 41%-50%; the fi fth grade-the rate of dry and red leaves was between 51%-60%; the sixth gradethe rate of dry and red leaves was between 61%-70%;the seventh grade-the rate of dry and red leaves was between 71%-80%; the eighth grade-the rate of dry and red leaves was between 81%-90%; and the ninth grade-the rate of dry and red leaves was≥90%.
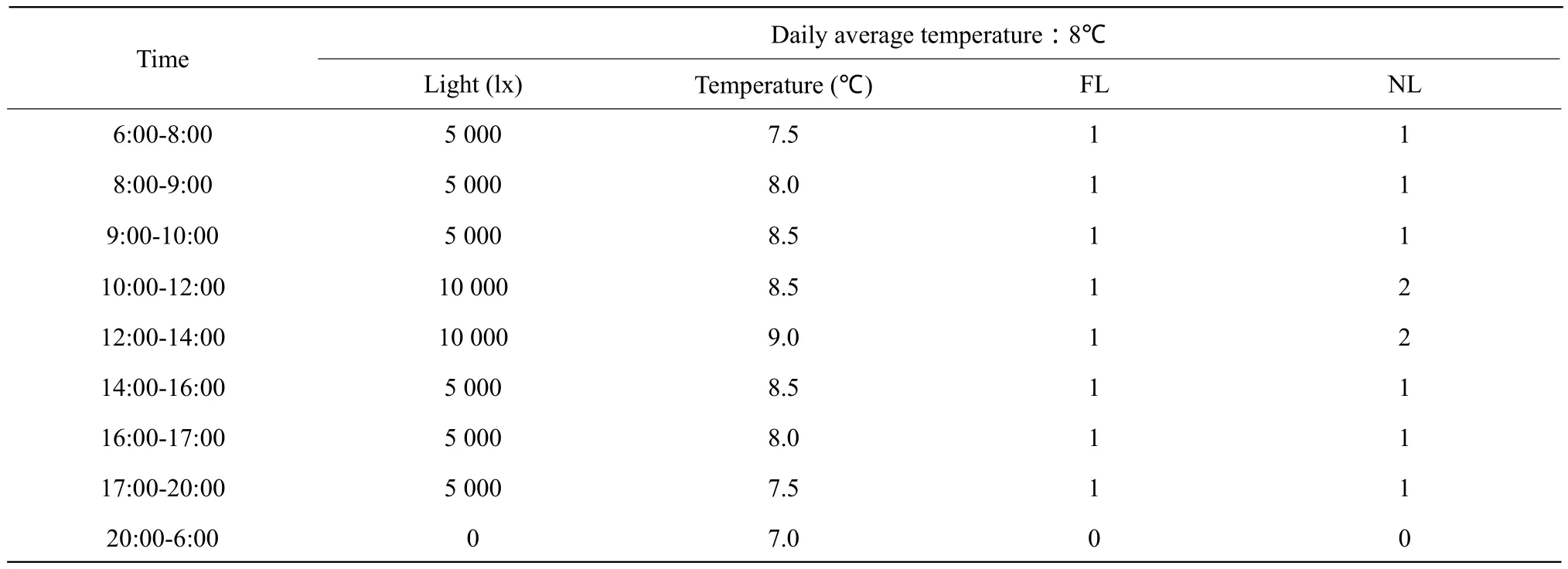
Table 1 Temperature and light setting for climate-control chamber

Fig. 1 Changes of natural air temperature
Determination of biomass
After the processing, selected 100 seedlings from each group respectively, rinsed them with distilled water,fi xed them at 105℃ for 15 min and then dried them to get constant dry weight of 100 seedlings (g).
Determination of the SPAD
The amount of chlorophyll content was measured by chlorophyll meter (SPAD-502 type). Gripped the center of the uppermost fully expanded leaves with chlorophyll meter to evaluate the SPAD value. Took 10 seedlings for one group and then got their average SPAD value. Repeated the process for three times.
Determination of net photosynthetic rate
Used LI-6400 portable photosynthesis system manufactured by American LI Company to determine the net photosynthetic rate of rice leaves (Pn).
Determination of chlorophyll fluorescence parameters
The chlorophyll fl uorescence parameters of rice leaves were determined by Germany WALZ PAM 2500 portable pulse modulated chlorophyll fl uorescence analyzer. Measurement methods referred to Genty et al.(1989). Took uppermost fully expanded leaves, made dark adaptation before the measurement for 20 min,and then opened the measuring light and got the original fl uorescence of F0, then opened the saturation pulse of light to get the maximum fluorescence Fm and maximal photochemical eff i ciency of Fv/Fm. Then opened the photochemistry, made light adaptation for 20 min and then got the fl uorescence parameters of the measured light existence of the maximum fl uorescence Fm', among which, Fv/Fm, qP, and qN were derived by the following formula:
Fv/Fm (maximum photochemical efficiency)=(Fm-F0)/Fm;
qP (variable photochemical devoid)=(Fm'-Ft)/(Fm'-F0);
qN was (non-photochemical fl uorescence quenching coeff i cient)=Fm/Fm'-1.
Determination of unsaturated fatty acids in leaves The oleic acid, linoleic acid and linolenic acid content were measured with reference to the methods by Yu et al. (1996). The formula was that the index of unsaturated fatty acids (UFA)=oleic acid content(mol%)×1+linoleic acid content (mol%)×2+linolenic acid content (mol%)×3.
Results
Effects of phosphate on cold tolerance ability of rice seedlings at low temperature
There was an obvious difference in cold resistance ability among different rice varieties (Table 2). Under low temperature conditions, with no application of phosphate, the rate of dry and red leaves of Jijing 81 was 17.6% and its resistance ability was the fi rst grade.In contrast, the rate of dry and red leaves of Changbai 9 was 31.2% and its resistance ability was the third grade. After adding phosphate to 0-75 g · m-2, the rate of dry and red leaves of both varieties declined but the rate of Jijing 81 reduced a little, Changbai 9 changed significantly. As for their cold tolerance abilities,Jijing 81 had no change and Changbai 9 increased up to grade 1. When phosphate was added to 100 g · m-2,the rate of dry and red leaves of both Jijing 81 and Changbai 9 increased, between which there was an obvious rise in Changbai 9 and its cold tolerance ability declined from grade 1 to grade 2.
Effects of phosphate on rice seedling biomass under low temperature condition
It could be seen from Fig. 2 that under normal temperature, the dry weight of 100 seedlings tended to increase fi rst and then decreased with the increase of phosphate amount. When the phosphate amount reached 50 g · m-2,both varieties reached their maximum in dry weight.While under low temperature condition, both dry weights were subject to varying degrees of inhibition of cold tolerance and the degree of inhibition of Jijing 81 was significantly less than that of non-cold tolerance variety Changbai 9. Compared with the dry wights, under room temperature without influence of phosphate, Jijing 81 and Changbai 9 declined by 19.12% and 27.8%. Although the trend was similar to that under room temperature, optimal application of phosphate had positive effect on rice seedlings. The dry weights of both varieties reached maximum when the amount of phosphate was 75 g · m-2.

Table 2 Cold tolerance identif i cation of rice at seedling stage under low temperature stress
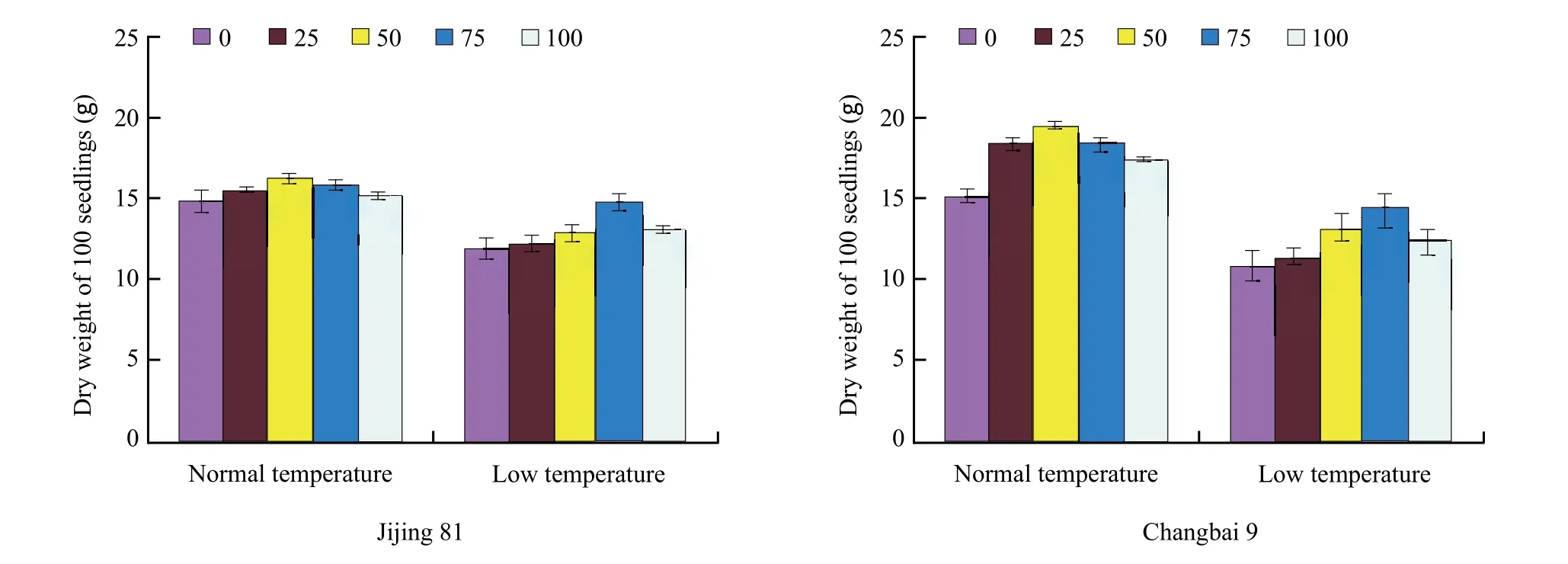
Fig. 2 Effects of phosphate fertilizer application on biomass in rice under low temperature stress
Effects of phosphate on the SPAD value of rice seedlings at low temperature
Table 3 showed that under normal temperature conditions, after applying phosphate, the SPAD values of both varieties were higher than that without the influence of phosphate except that the phosphate application amount was 25 g · m-2and their SPAD value reached their maximum at the phosphate level of 50 g · m-2. If more phosphate were used, their SPAD value showed a downward trend. While under low temperature conditions, the SPAD values of both seedlings leaves decreased greatly and different cold tolerant varieties showed different values. In contrast to the results in room temperature, without the application of phosphate at low temperature, the SPAD of cold tolerance variety Jijing 81 decreased by 5.97% and non-cold-tolerance Changbai 9 decreased by 14.5%. With the increase of phosphate application,their SPAD values did not decrease so much as before.When the phosphate level was for 75 g · m-2, the SPAD values of Jijing 81 and Changbai 9 leaf declined only 2.9% and 4.2%, respectively.
Effects of phosphate on net photosynthetic rate of rice seedlings under low temperature stress
It could be seen from Fig. 3 that at room temperature,the net photosynthetic rate of Jijing 81 and Changbai 9 tended to increase fi rst and then decreased with the increased in phosphate levels, and both reached their maximum when the phosphorus amount was 50 g · m-2. At low temperature, the net photosynthetic rate of Jijing 81 and Changbai 9 decreased signifi cantly, in which the rate of Jijing 81 was higher than that of Changbai 9. When the phosphate amount was 25-75 g · m-2, the net photosynthetic rate of the two varieties increased and reached their maximum at the phosphate level of 75 g · m-2. When the phosphate amount was 100 g · m-2, their net photosynthetic rates decreased and the rate of the cold-resistance variety Changbai 9 decreased more than that of Jijing 81.
Effects of phosphate on fluorescence parameters of rice seedlings under low temperature conditions
Table 4 showed that at room temperature, with the increase of phosphate levels, there was a slight decrease in Fv/Fm of the two varieties but no significant difference under different P treatments.Under low temperature, compared with room temperature, Fv/Fm of the two varieties decreased signif i cantly in which Fv/Fm of Jijing 81 was always higher than that of the cold non-resistant variety Changbai 9. When the phosphate level was 25-75 g · m-2,Fv/Fm of the two varieties increased with increasing phosphate levels. When the phosphate amount was 75 g · m-2, Fv/Fm of the two varieties increased by 11.7%and 24.3%, respectively. If more phosphate was added,Fv/Fm of Changbai 9 showed a significant decrease and that of Jijing 81 had no big decline. It also could be seen from Table 4 that at room temperature, qP of Jijing 81 and Changbai 9 had no signif i cant change no matter phosphate was added or not. While under low temperature stress, qP of both varieties decreased. But with the continual increase of phosphate, qP of the two did not decrease so much as before and reached their minimum decrease at the phosphate level of 75 g · m-2.If more phosphate was added, qP of the two varieties decreased signif i cantly.

Table 3 Effects of phosphate fertilizer application on photosynthesis in rice under low temperature stress

Fig. 3 Effects of phosphate fertilizer application on photosynthesis in rice under low temperature stress

Table 4 Effects of phosphate fertilizer application on photosynthesis in rice under low temperature stress
At normal temperature, with the increase in phosphate level, qN of Jijing 81 and Changbai 9 decreased and had little change among the treatments; while in low temperature conditions, there was obvious difference among different chilling tolerance varieties.Compared with the number obtained at normal temperature, qN of Jijing 81 increased signif i cantly while qN of cold tolerance variety Changbai 9 decreased a bit at the phosphate level of 0-25 g · m-2.
Effects of phosphate on unsaturated fatty acid in rice under low temperature stress
It can be seen from Table 5 and Fig. 4 that under normal temperature conditions, with the increase of phosphate level, the oleic acid, linoleic acid and linolenic acid in the leaves of Jijing 81 and Changbai 9 increased slightly and there was no big difference among treatments; saturated fatty acid index had no signif i cant change. Compared with non-cold-resistance variety Changbai 9, the leaves of cold-tolerance variety Jijing 81 contained more unsaturated fatty acid. At low temperature compared with room temperature, oleic acid content in the leaves of Jijing 81 decreased but its linoleic acid and linolenic acid content increased;in contrast, oleic acid and linoleic acid content in the leaves of Changbai 9 decreased, but fl ax acid content increased. If the phosphate level reached 25-75 g · m-2,oleic acid content in the leaves of both varieties did not change signif i cantly, linoleic acid content reduced and unsaturated fatty acids signif i cantly increased.

Table 5 Effects of phosphate fertilizer application on unsaturated fatty acid (mol%)in rice under low temperature stress

Fig. 4 Effects of phosphate fertilizer application on index of unsaturated fatty acid in rice under low temperature stress
Discussion
Japan started early on the study of rice chilling injury(Xu et al., 2000). China began such study since 1970s to 1980s. Combined with various rice cold damage characteristics in different areas, we developed corresponding identification and evaluation methods on cold tolerance ability of rice. At present, the main evaluation method is to determine the proportion of red and dry leaves (Han et al., 2004; Li et al., 2006).This study identified the cold tolerance abilities of rice seedlings which were added with phosphate under low temperature conditions. The results showed that different rice varieties differred greatly in their abilities to resist cold. Appropriate amount of phosphate could improve the cold tolerance capacity of rice seedlings and enhanced their cold tolerance levels. Chillingsensitive tolerance variety was more sensitive to the increase in phosphorus.
Phosphorus plays an important role in cells and the metabolic process and is closely linked to protein synthesis, cell division and cell growth. Studies have shown that phosphorus deficiency will inhibit the formation and proliferation of cells, resulting in the growth and development stagnation (Li et al.,2007). The low temperature would also inhibit the growth of the plants. When rice seedlings encounter low temperature stress, the biological characteristics of the plant such as fresh weight and dry weight are significantly affected, and there was an obvious difference among different varieties (Li et al., 2006).This study showed that regardless of room temperature or low temperature conditions, phosphorus contributed to the increase of dry weight of rice seedlings per hundred. However, at low temperature, cold-resistant varieties were less inhibited than non-coldresistant varieties. The fact that the optimal phosphate dosage of the two varieties had been improved signif i cantly indicated that the appropriate amount of the phosphorus could enhance the quality of the rice seedlings and increase rice biomass to a certain extent and alleviated the low temperature damage to the seedlings. Results were consistent with the study on watermelon (Chen et al., 2007)and tomato (Li et al.,2007).
Photosynthesis in plants is sensitive to low temperature. Studies have shown that short-term low temperature will result in the accumulation of phosphorylated intermediates and thus lower phosphorus levels in the cytoplasm (Hurry et al., 1998)and inhibit the synthesis of the chloroplast (Zeng et al., 2000). The content of chloroplast is at certain degree linked to photosynthesis (Zhang et al., 2000). The results of this study showed that regardless of room temperature and low temperature, phosphorus contributed to the improvement of relative chlorophyll content and net photosynthetic rate in rice seedling leaves, but excessive phosphorus would lead to lower chlorophyll content and net photosynthetic rate. At low temperatures, cold-resistant varieties contained less relative chlorophyll content and the net photosynthetic rate decreased, but they were always higher than that of the chilling-sensitive tolerance varieties. The optimal temperature for phosphate was room temperature, but under low temperature, proper increased in P fertilizer to a certain extent slowed the decrease in relative chlorophyll content of rice seedlings and maintained a relatively higher net photosynthetic rate of leaves.
The study found that Fv/Fm was sensitive indicator for the cold tolerance capacities of plants.Under stressful conditions, the value would significantly reduce (Zhao et al., 2000). Under low temperature stress, cold hardening exercised cucumber seedlings has a higher qN value than those without any exercises (Chen et al., 1995). Wang (2005)found that under low temperature stress, cold-resistant varieties would produce higher qP and qN to dissipate excessive light energy to reduce excessive excitation energy accumulation in their reaction centers and lower the impact on PSⅡ reaction center and antenna system to protect photosynthetic apparatus. While as for the cold-sensitive varieties, their reaction centers and antenna systems are seriously affected under low temperature stress, that is qP decreased quickly, but qN decreased with the extension of low-temperature time, resulting in decreased photosynthetic rate.Low P can reduce the PS II light energy conversion and electron transfer efficiency in leaves and thus increase excessive excitation energy (Li et al.,2004). The results of this study showed that Fv/Fm,qP and qN of different chilling-tolerance varieties at room temperature did not change significantly with phosphate increase; while at low temperature,Fv/Fm and qP, qN of the two varieties decreased,indicating that low temperatures led to the inhabitation of PSⅡ photochemical activity and the damage of the photosynthetic apparatus which resulted in the reduction of PSⅡ light energy conversion and electron transfer eff i ciency in leaves. Therefore, under low temperature, appropriate increase in P fertilizer could improve eff i ciency of light conversion for both chilling-tolerance and non-cold-resistant varieties.Meanwhile, the higher qP could cause fluorescence quenching and reduce the damage of excess excitation energy to reaction center and reduce heat loss.
Numerous studies showed that the unsaturated fatty acids with increased membrane lipid could effectively reduce the phase transition temperature of the membrane lipid. The greater the proportion of unsaturated fatty acid composition of membrane lipid and the lower phase transition temperature was, the stronger its resistance to cold would be (Shen et al., 1997).Meanwhile, the cold resistance plants generally have higher membrane lipid saturation which helps to maintain liquidity and normal physiological functions at a lower temperature. The results of this study showed that regardless of room temperature or low temperature, cold-resistant rice varieties had higher unsaturated fatty acid index and hence contained more unsaturated fatty acid. The increase of phosphate at room temperature had few effects on the unsaturated fatty acid index of different cold tolerant rice varieties;the unsaturated index of chilling-tolerance seedlings was signif i cantly higher than that of chilling-sensitive tolerance. In contrast, the increase in the amount of phosphorus at low temperatures, the unsaturated fatty acids index of both cold tolerance and chillingsensitive tolerance varieties were significantly increased, indicating that increasing P fertilizer at low temperature could increase unsaturated fatty acids and fasten the fl ow of membrane lipid, thereby reduce the harm to the rice caused by the low temperature.
Conclusions
Phosphorus played a very important role in enhancing the cold tolerance capacity of rice seedlings under low temperature. Appropriate increase in P fertilizer in the nursery bed soil could enhance the level of cold tolerance ability of rice seedlings under low temperature and improve their qualities, slow down and reduce the degree of relative chlorophyll content of rice seedlings, improve the net photosynthetic rate,photochemical efficiency and unsaturated fatty acid index. The increased phosphorus had more signif i cant effects on chilling-sensitive tolerance rice than on chilling-tolerance rice variety. At present, the average phosphorus content in seedlings nursery bed soil was about 50 g · m-2in the eastern part of Jilin Province.In order to prevent cold damage, attention should be paid not only on rice variety, but also on changing the proportion of phosphorus in seedling bed soil. The results of this study indicated that a phosphate level in about 75 g · m-2could effectively improve rice tolerance to cold and reduce the damage degree.
Chen Y Z, Li X P, Xia L. 1995. The application of chlorophyll fl uorescence technique in the study of respones of plants to environmental stresses. Journal of Tropical and Subtropical Botany, 3(4):79-86.
Chen G, Zhou M B, Tan S T. 2007. Effect of phosphorus, boron nutrition on physiological and biochemical characteristics of watermelon seedlings under low temperature. Journal of Fruit Science, 24(6): 815-819.
Genty B E, Briantais M J, Baker N R. 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fl uorescence. Biochimica and Biophysica Acta, 990:87-92.
Han L Z, Zhang S Y. 2004. Methods of characterization and evaluation of cold tolerance in rice. Journal of Plant Genetic Resources, 5(2):75-80.
Hurry V, Huner N, Selstam E, et al. 1998. Photosynthesis at low temperature. In: Raghavendra A S. Photosynthesis, a comprehensive treatise. Cambridge University Press, Cambridge. pp. 87-101.
Li S Y. 2004. The effect of Mo, P compound on the growth and coldresistance of winter wheat. Huazhong Agricultural University,Wuhan.
Li S Z, Hu C H, Gong J. 2004. Effects of low phosphorus stress on the chlorophyll fl uorescence of different phosphorus use eff i cient maize(Zea mays L.). Acta Agronomica Sinica, 30(4): 365-370.
Li X, Dai C C, Cheng R. 2006. Identification for cold tolerance at different growth stages in rice (Oryza sativa L.)and physiological mechanism of differential cold tolerance. Acta Agronomica Sinica,32(1): 76-83.
Li Xi. 2007. Effects of low temperature stress of phosphate fertilizer on heliogreenhouse tomato seedlings growth and physiological activity.Acta Agriculturae Boreali-Sinica, 22(5): 142-146.
Shen M, Wang M X, Huang M R. 1997. Advances in research on chilling resistance mechanisms of plants. Chinese Bulletin of Botany,14(2): 1-8.
Wei X H, Li H X, Lu S N. 2005. Study on the effect of the different ration of nitrogen, phosphorus and potassium on yields of sugarcane by the optimization compound desig. Soils and Fertilizers, 4: 6-10.
Wang D M, Zhou J H, Zhu X L. 1999. Effect of P, Ca, Zn nutrition of growth of flue-cured tobacco andlts resistance to unfavorable environmental conditions. Acta Tabacaria Sinica, 5(3): 23-27.
Wang G L, Guo Z F. 2005. Effects of chilling stress on photosynthetic rate and the parameters of chlorophyll fluorescence in two rice varieties differing in sensitivity. Chinese Journal of Rice Science,19(4): 381-383.
Wang C Y. 2008. Northeast crops chilling damage study. Meteorological Press, Beijing.
Xu F R, Dai L Y, Ye C R. 2000. Research status of tolerance to cold in rice. Crops, 1: 4-5.
Yu X J, Liu Y Y, Hu J B. 2003. Studies on climate change in Jilin province in recent 50 years. Jilin Meteorological, 2: 27-30.
Yu H G, Su W A. 1996. Studies on relationship between fatty acids desaturation of PS II membrane and low temperature photoinhibition of cucumber. Acta Biophysica Sinica, 11(2): 227-233.
Zeng N Y, He J X, Zhao W. 2000. Changes in components of pigments and proteins in rice photosynthetic membrane during chilling stress.Acta Botanica Boreali-Occidentalia Sinica, 20(1): 8-14.
Zhang G M, Wang L M, Wang L Z. 2000. The effect of low temperature on chlorophyll content and growth of maize at seedling stage.Heilongjiang Agricultural Sciences, 1: 10-12.
Zhao H J, Zou Q, Yu Z W. 2000. Chlorophyll fluoresence analysis technique and its application to photosynthesis of plant. Journal of Henan Agricultural University, 34(3): 248-251.
杂志排行

Journal of Northeast Agricultural University(English Edition)的其它文章
- Establishment of TaqMan Real-time Quantitative PCR Assay for Foreign Gene Copy Numbers in Transgenic Soybean
- Study on Semen Freezing Preservation of German Shepherd Dogs
- In Vitro Study on Commercial Organic Acid Activate WD (WD)Against Four Pathogenic Bacteria
- Response of Osmotic Adjustment of Lactobacillus bulgaricus to NaCl Stress
- Temperature Simulation of Berry Slices Under Microwave Vacuum Puff i ng Conditions
- Storage System Design Scheme in Virtualization Construction