传染性法氏囊病病毒NN1107广西株的分离鉴定及遗传进化分析
2012-06-29兰美益陈泽祥韦志锋秦若甫
李 军,陶 立,兰美益,陈泽祥,韦志锋,秦若甫,彭 昊,杨 威*
(1.广西兽医研究所,广西 南宁 530001;2.广西畜禽疫苗新技术重点实验室,广西 南宁 530001)
传染性法氏囊病病毒(Infectious bursal disease virus,IBDV)是鸡传染性法氏囊病(Infectious bursal disease,IBD)的病原体。IBDV感染幼龄鸡后,主要在法氏囊髓质区的未成熟B淋巴细胞和B淋巴细胞前体细胞中增殖,引起感染细胞的变性和坏死,临床上出现以法氏囊水肿、出血和萎缩为特征的病理变化[1-2]。IBDV在鸡群中的传播不仅导致易感鸡群的大量死亡,IBDV的杀淋巴细胞作用还能引起鸡群的免疫应答能力降低,使鸡群对其他病毒或细菌感染的敏感性增加,给养鸡业造成巨大的经济损失[3-4]。近十几年来,随着IBDV疫苗的广泛应用,IBDV在疫苗免疫选择压力下,自身不断发生变异,毒力获得增强,出现经典毒株(classical IBDV,cIBDV)向超强毒株(very virulent IBDV,vvIBDV)的演化,vvIBDV可以突破高水平的母源抗体保护,导致鸡发病年龄提前和病死率增高,给IBD的防控带来了困难。
IBDV属于双RNA病毒科、禽双RNA病毒属成员,基因组由A、B两个双链RNA片段组成[5]。A片段的VP2基因是IBDV主要结构蛋白VP2的编码基因,VP2蛋白位于病毒粒子的外表面,是IBDV的主要毒力因子,能诱导细胞凋亡和诱导产生保护性中和抗体[6]。研究证实VP2序列中的VP2高变区(vVP2)与IBDV的毒力密切相关。vVP2位于VP2蛋白的第206~350位氨基酸之间,由大小两个亲水区和一个七肽区组成,构成一个构象依赖型的抗原表位,诱导机体产生中和抗体。vVP2内氨基酸的改变会引起IBDV 毒力的改变[7-9]。
阳秀英等和何秀苗等2000年—2007年对广西鸡群IBDV的分子流行病学研究表明,广西鸡群中主要流行vvIBDV,各地毒株来源复杂,部分IBDV毒株的抗原性可能发生漂移[10-11]。2011年7月,广西南宁市某养鸡场饲养的23日龄三黄鸡暴发了一起IBD,病死率为6%。本研究对病死鸡进行了IBDV分离和鉴定,并对其vVP2进行克隆和测序,将获得的vVP2序列与广西流行的IBDV毒株vVP2序列进行比较分析,建立遗传进化树,从分子水平上对广西近期IBDV的变异情况进行追踪和分析。
1 材料与方法
1.1 材料
1.1.1 病料 来自广西南宁市某养鸡场的临床疑似IBD的病死鸡,采其法氏囊作为病料。
1.1.2 鸡胚 9日龄健康鸡胚,购自广西华桂源种禽有限公司。
1.1.3 试剂和仪器 PCR Taq mix、DNA Marker 1、DNA胶回收试剂盒、质粒小量抽提试剂盒购自广州东盛生物科技有限公司;病毒基因组DNA/RNA提取试剂盒购自天根生化科技(北京)有限公司;pMD18-T载体、RNA酶抑制剂和 MLV反转录酶购自宝生物工程(大连)有限公司;PCR仪购自日本TaKaRa公司。
1.2 方法
1.2.1 病毒分离 病毒的分离方法按阳秀英等的方法进行[10]。
1.2.2 病毒分离株RNA的提取 用病毒基因组DNA/RNA提取试剂盒提取接种病毒的鸡胚尿囊液中的总RNA,方法按说明书进行。
1.2.3 病毒分离株vVP2基因的扩增、克隆和测序按韦平等的方法进行IBDV的RT-PCR检测[12]。利用DNA胶回收试剂盒回收PCR扩增产物。回收纯化后与pMD-18T载体连接,转化大肠埃希菌DH5α,用质粒小量抽提试剂盒抽提质粒,经双酶切和PCR鉴定后,将阳性重组质粒送上海英骏生物技术有限公司测序。
1.2.4 分离株vVP2序列的遗传进化分析 应用DNA Star软件包的MegAlign软件对测序获得的vVP2片段序列与GenBank登录的5株IBDV疫苗株和26株2000年—2010年在广西流行的IBDV野毒株的vVP2序列进行核苷酸和氨基酸的序列比较和同源性分析,绘制遗传进化树。分析中应用的毒株信息见表1。
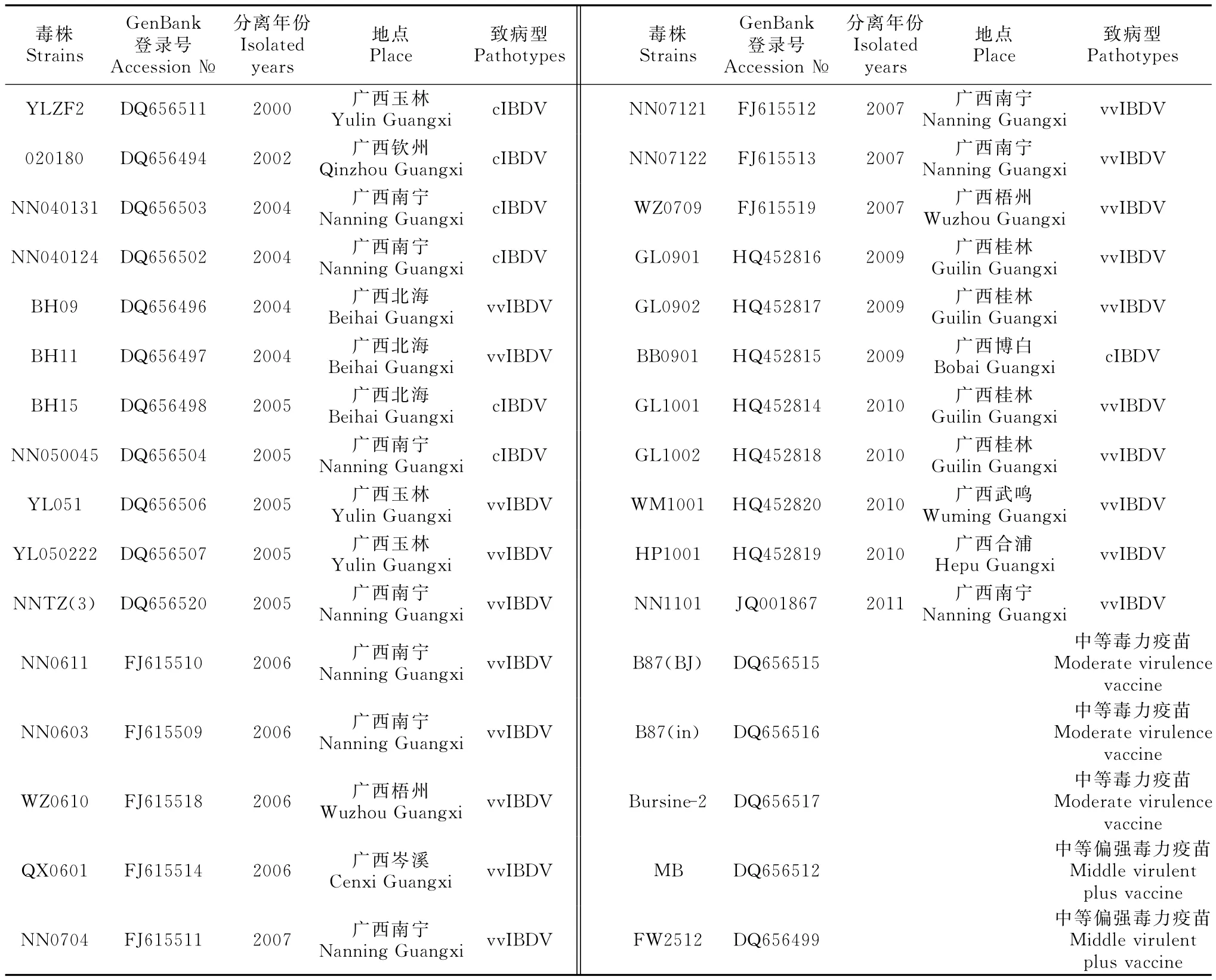
表1 所用的IBDV毒株信息Table1 The information of IBDV strains used in this study
2 结果
2.1 病毒分离和鉴定
发病鸡的法氏囊研磨过滤后经绒毛尿囊膜途径接种9日龄健康鸡胚,鸡胚在43h后死亡,胚体表现为水肿和出血。取尿囊液进行血凝试验和细菌培养检测均为阴性。应用韦平等[12]的方法对鸡胚尿囊液进行IBDV的RT-PCR检测,扩增出471bp的目的片段,证实从病料中分离到了IBDV,将此毒株命名为NN1107。
2.2 分离株vVP2序列的比较分析
2.2.1 核苷酸序列同源性比较 对NN1107分离株的vVP2进行克隆和测序,获得了NN1107分离株vVP2核苷酸序列,长度为471bp。对序列进行限制性内切酶酶切位点检测,在271位核苷酸处有一个vvIBDV的特征性SspI酶切位点。将此序列与广西野毒株和疫苗株的核苷酸序列进行同源性比较,NN11017株与广西vvIBDV毒株的同源性在96%~99.6%之间,其中与野毒株 NN07122和HP1001的同源性最高,为99.6%。NN1107株与广西cIBDV毒株的同源性在90.3%~91.6%之间;与疫苗株的同源性则在90.3%~96.6%之间。
2.2.2 氨基酸序列同源性比较 将推导的氨基酸序列与广西野毒株和疫苗株的氨基酸序列进行同源性比较,NN1107株与广西vvIBDV毒株的同源性在94.9%~99.4%之间,其中与野毒株 BH09、NNTZ(3)、NN07122和 HP1001的同源性最高,为99.4%。NN1107株与广西cIBDV毒株的同源性在91.1%~93%之间;而与疫苗株的同源性在91.8%~97.5%之间,其中与 B87(in)、Bursine-2、FW2512株的同源性均为91.8%;与 B87(BJ)和MB株的同源性则为93%和97.5%。
2.2.3 氨基酸序列分析 NN1107株vVP2氨基酸序列中与病毒毒力和抗原相关的氨基酸均符合vvIBDV 特 征[13-14],即第326~332位的七肽区SWSASGS序列保持不变;大亲水一区(第212~224位氨基酸)内212、213、214和222位氨基酸为N、D、Y和A;大亲水二区(第314~324位氨基酸)内318、321、323和324位氨基酸为G、G、D和Q;小亲水一区(第248~252位氨基酸)内249位氨基酸为Q;小亲水二区(第279~290位氨基酸)279和284位氨基酸为D和T。其他与病毒毒力和抗原相关的氨基酸位点,如第242、253、254、256、270、294、299和330位氨基酸分别为I、Q、G、I、A、I、S和S。NN1107株与广西部分野毒株和疫苗株的vVP2氨基酸序列比较见图1。
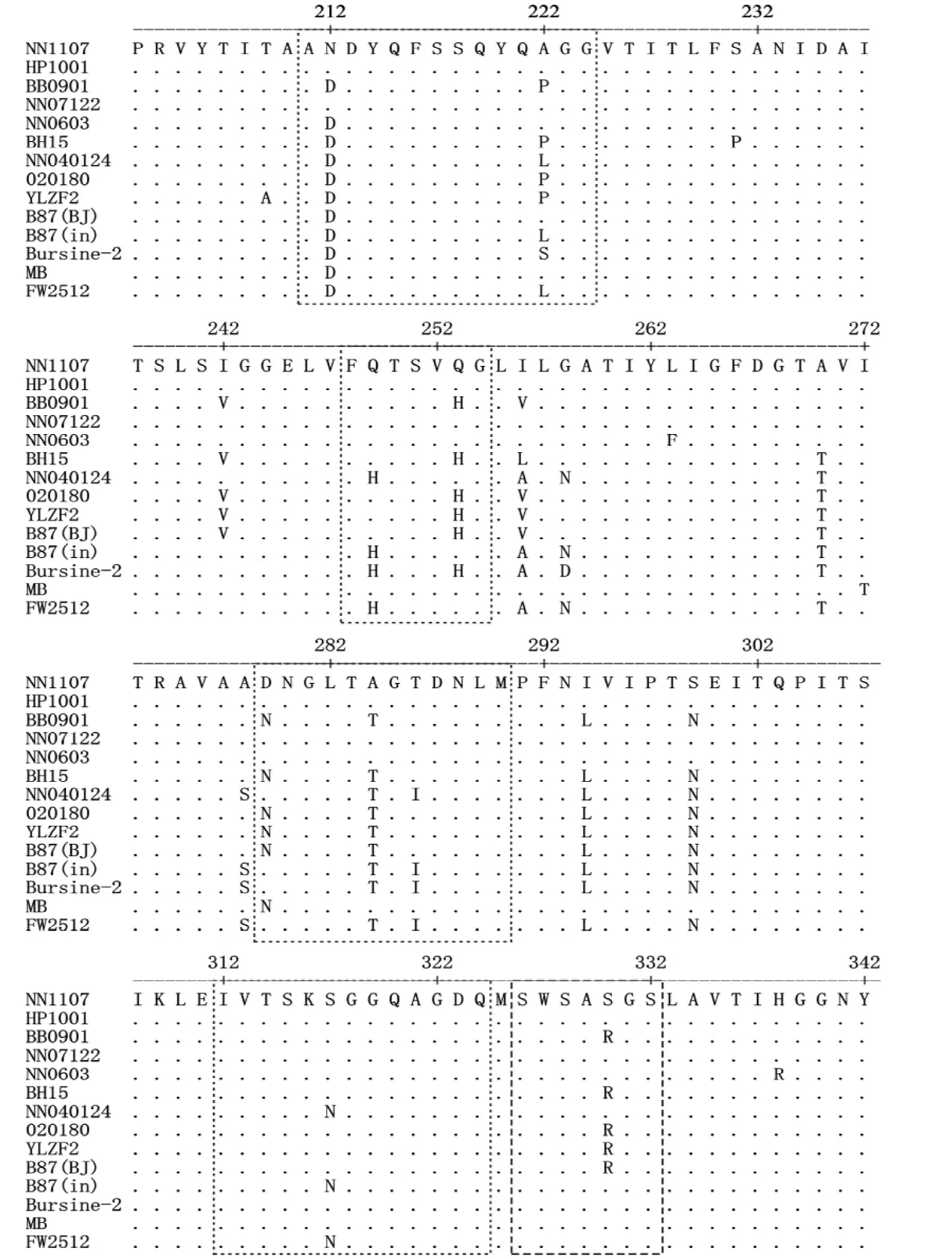
图1 vVP2的氨基酸序列比较Fig.1 Comparison of the amino acid sequences of vVP2
2.3 分离株vVP2序列的遗传进化分析
将NN1107株与5株IBDV疫苗株和26株2000年—2010年在广西流行的IBDV野毒株的vVP2核苷酸序列进行同源性比较,绘制遗传进化树(图2)。可以发现遗传进化树由vvIBDV毒株、cIBDV毒株和疫苗株3个群组成,群间的亲缘关系较远。NN1001株属于vvIBDV毒株群,它与曾经在2004年、2005年、2007年和2010年流行的毒株BH09、NNTZ(3)、NN07122、HP1001的亲缘关系最近,共同组成亚群2;而2006年、2007年和2010年的大部分毒株以及2004年至2005年流行的大部分毒株分别组成亚群1和亚群3。
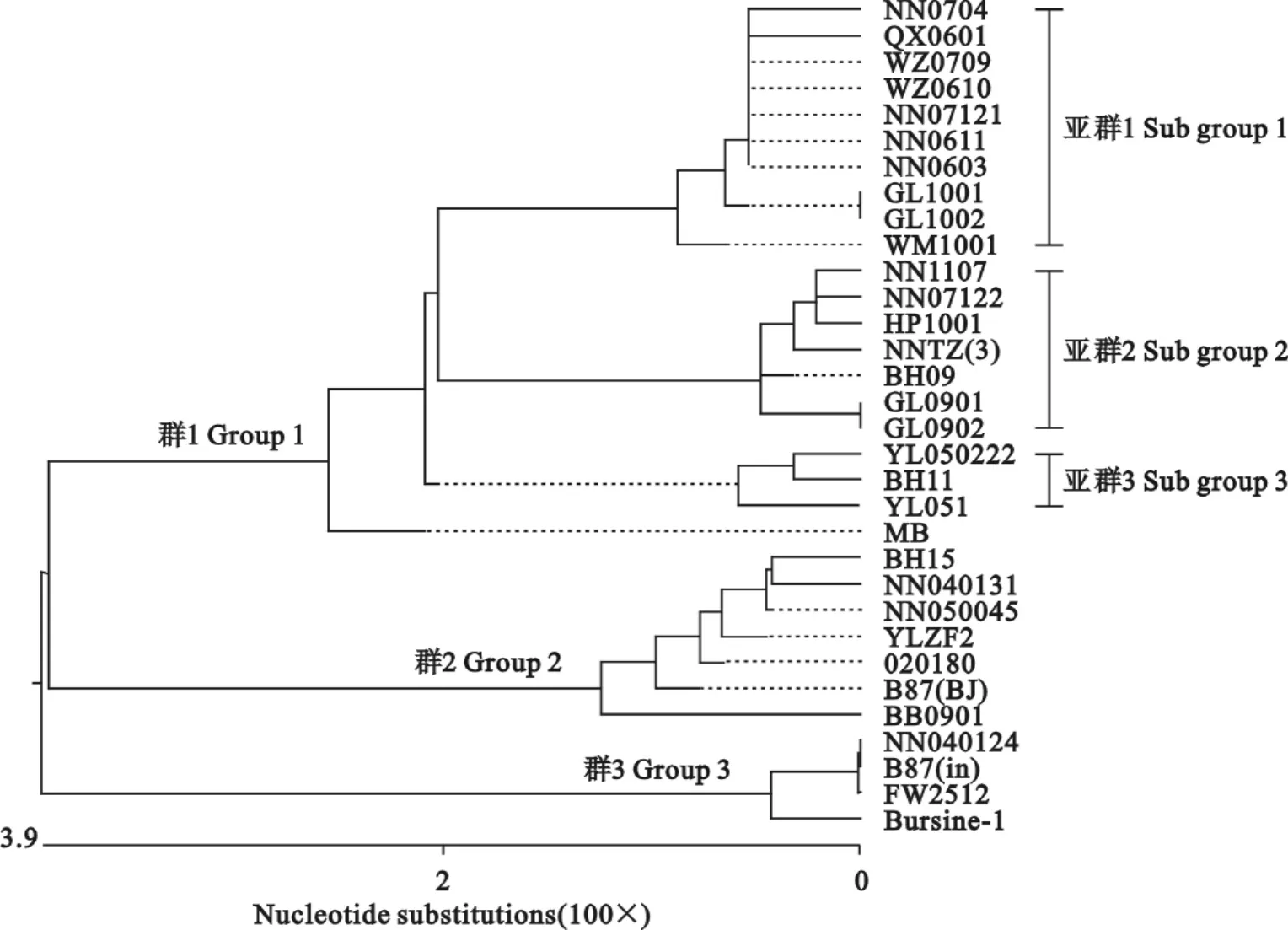
图2 根据vVP2核苷酸序列绘制的遗传进化树Fig.2 Phylogenetic tree of isolate and reference strains based on the nucleotide sequences of vVP2
3 讨论
采用鸡胚绒毛尿囊膜接种和RT-PCR检测技术对临床疑似IBD的病死鸡法氏囊进行IBDV的分离和鉴定,成功分离出一株IBDV(NN1107株)。对NN1107分离株vVP2片段进行克隆和测序,序列分析和同源性比较结果显示,NN1107株具有vvIBDV的特征性酶切位点SspI,且与广西vvIBDV野毒株的同源性极高,表明NN1107株为一株vvIBDV。
VP2蛋白不仅是IBDV的一个主要结构蛋白,诱导机体产生保护性中和抗体,还是一个变异最大的蛋白,其变异主要集中在vVP2区域(第206~350位氨基酸),vVP2内氨基酸的改变都有可能使病毒的毒力增强,突破疫苗的免疫保护[7-9]。因此,可以通过vVP2序列的同源性比较来分析毒株的遗传进化。vVP2氨基酸序列同源性比较和遗传进化分析显示,在近10年,广西同时存在着cIBDV和vvIBDV两种不同毒力的IBDV毒株流行。本研究分析的27株IBDV毒株中,只有7株毒株属于cIBDV,且毒株大都分离于2005年以前,只有1株BB0901是分离于2009年;而vvIBDV毒株分离自2004年至2011年,表明广西目前IBDV的主要流行毒株仍然为vvIBDV。
NN1107株与2004年毒株BH09、2005年毒株NNTZ(3)、2007年毒株 NN07122和2010年毒株HP1001的同源性高达99.4%;其主要氨基酸位点也未发生任何改变,说明这些毒株很有可能来源于同一病毒株的进化。本研究构建的遗传进化树也显示vvIBDV在进化中也分成2个亚群,在同一地区,如位于桂南地区的南宁市和位于桂北地区的桂林市都存在有不同亚群的vvIBDV,表明广西vvIBDV来源复杂,流行不受地域限制。
从vVP2核苷酸序列绘制的遗传进化树可以看出,NN1107株等vvIBDV毒株与传统的疫苗株B87(in)、Bursine-2、FW2512株分属于两个基因群。在与病毒毒力和抗原相关的氨基酸位点中,有10个位点,即第212、222、249、256、270、279、284、286、294和299位的氨基酸发生了改变。这些氨基酸的改变可能会导致中和抗体识别的抗原表位发生构象上的变化,使疫苗株 B87(in)、Bursine-2、FW2512株诱导机体产生的中和抗体不能有效地与病毒结合,阻断vvIBDV感染B淋巴细胞,这可能是vvIBDV突破高水平母源抗体保护的原因。
[1]Lukert P D,Saif Y M.Infectious Bursal Disease[M].In:Calnek B W,Barnes H J,Beard C W,et al(Eds),Diseases of Poultry(10th ed.).Iowa State Universuty Press,1997:721-738.
[2]毛炳宇,马晓丽,赵 晖,等.人工感染IBDV鸡法氏囊的电镜研究 [J].畜牧兽医学报,1998,29(5):455-461.
[3]Imai K,Mase M,Tsukamoto K,et al.Persistent infection with chicken anaemia virus and some effects of highly virulent infectious bursal disease virus infection on its persistency[J].Res Vet Sci,1999,67(3):233-238.
[4]Bautista D A,Elankumaran S,Heckert R A.Effect of a variant infectious bursal disease virus(E/Del)on Salmonella typhimuriuminfection in commercial broiler chickens[J].Avian Dis,2004,48(2):361-369.
[5]Hudson P J,McKern N M,Power B E,et al.Genomic structure of the large RNA segment of infectious bursal disease virus[J].Nucleic Acids Res,1986,14:5001-5012.
[6]Fernandez-Arias A,Martinez S,Rodriguez J F.The major antigenic protein of infectious bursal disease virus,VP2,is an apoptotic inducer[J].J Virol,1997,71:8014-8018.
[7]Bayliss C D,Spies K U,Shaw R W,et al.A comparison ofthe sequences of segment A of four infectious bursal disease virus strains and identification of a variable region in VP2[J].J Gen Virol,1990,71(Pt6):1303-1312.
[8]Schnitzler D,Bernstein F,Müller H,et al.The genetic basis for the antigenicity of the VP2protein of the infectious bursal disease virus[J].J Gen Viro,1993,74(Pt 8):1563-1571.
[9]Eterradossi N,Arnauld C,Toquin D,et al.Critical amino acid changes in VP2variable domain are associated with typical and atypical antigenicity in very virulent infectious bursal disease viruses[J].Arch Virol,1998,143(8):1627-1636.
[10]阳秀英,韦 平,黄志永,等.我国部分省区鸡传染性法氏囊病病毒的分子流行病学研究 [J].畜牧兽医学报,2007,38(6):581-588.
[11]何秀苗,韦 平,官丁明,等.2000~2007年广西鸡传染性法氏囊病病毒的分子流行病学 [J].病毒学报,2009,25(6):437-444.
[12]韦 平,龙进学,阳秀英,等.传染性法氏囊病病毒快速检测与分型技术的研究 [J].中国兽医学报,2004,24(4):313-316.
[13]许信刚,李健强,王笑梅,等.传染性法氏囊病病毒超强毒HZ株和XN株VP2基因的克隆和序列比较分析 [J].动物医学进展,2000,21(1):39-42.
[14]曹永长,毕英佐,梁志清,等.超强传染性法氏囊病病毒宿主保护抗原的分子特征 [J].中国兽医学报,1998,18(6):521-526.
