楸树无性系表型识别特征与遗传变异1)
2012-06-13贠慧玲王军辉
贠慧玲 王军辉
(甘肃省小陇山林业科学研究所,天水,741022) (国家林业局林木培育重点实验室(中国林业科学研究院林业研究所))
张宋智 马建伟 赵秋玲 马丽娜
(甘肃省小陇山林业科学研究所)
皮孔是植物经历了长期自然选择,经过适应不同的自然环境而留下来的重要生理器官,皮孔的数量和形态是植物种鉴定的重要依据,也和植物的抗性有直接的关系[1-5]。植物皮孔是植物体与外界环境进行水分交换的通道,对植物的光合、呼吸、蒸腾等生理活动起着重要的调节作用[6-9]。皮孔的形状、大小、分布以及皮孔指标等还直接影响植物对水肥的吸收。以往研究表明,在寄主的抗病因素中,皮孔是病菌侵入的主要途径和潜伏的主要部位[10],皮孔密度及大小与病害发生有密切关系[11]。而有关楸树(Catalp bungeiC.A.Mey)皮孔的研究尚未见报道。本研究主要探讨楸树无性系皮孔的识别特征,对其不同无性系间皮孔大小、密度及其变异范围进行分类,初步了解各个无性系的特点,为楸树无性系的早期选择提供一定的理论依据。
1 材料与方法
供试材料由中国林科院林业所领导的国家楸树育种协作组提供的1年生楸树杂种无性系嫁接苗,2006年3月造林,共29个无性系。试验设计为随机完全区组设计,4株小区,4次重复。株行距为2.0 m×1.5 m。到2010年林龄为5年生树种。
皮孔性状的测定:在2010年7月的生长旺盛期,选择第2和第3次重复,每个无性系测量4个分生株,在树干距地1 m的高度向上,每个分生株测量3个样方(每个样方为5 cm2),每个无性系共测量12个样方。用游标卡尺测量每个样方上的每个皮孔的长、宽,并计算皮孔面积和皮孔数量。皮孔面积公式为S1=a1×b1×0.785 4;其中,S为皮孔面积,a为皮孔长,b为皮孔宽,0.785 4为系数。皮孔密度为单位面积上的皮孔数量,并用微距拍照。
叶痕性状的测定:对测定皮孔的每个分生株,测定树干中部1 m段的叶痕数量。抽样中部树干叶痕1轮,用游标卡尺测定叶痕长、叶痕宽,计算叶痕的面积和数量。叶痕面积公式:S2=a2×b2×0.785 4;其中,S为叶痕面积,a为叶痕长,b为叶痕宽,0.785 4为系数。同时调查树皮开裂程度及开裂方式,并用微距拍照。
生长性状的测定:2010年11月测定各无性系的树高和胸径。
数据统计以小区平均值参与数据处理,用Excel 2003和SPSS16.0进行统计分析。
2 结果与分析
2.1 皮孔性状的变异
2.1.1 楸树无性系皮孔密度
对29个无性系皮孔密度进行方差分析表明,不同无性系间呈极显著性差异,见表1。

表1 楸树无性系皮孔性状的遗传参数
皮孔密度最大的无性系为002-1(5.24个/cm2),皮孔密度最小的无性系2-1(1.80个/cm2)。对皮孔密度聚类分析,从大到小可以分为3类,见表2。第1类密集型(m≥4.26个/cm2);第2类较密集型(3.62个/cm2≤m≤3.94个/cm2);第3类疏散型(m≤3.3个/cm2)。皮孔密度平均为2.836个/cm2,变幅为 1.80 ~5.24 个/cm2,遗传变异系数为28.3%。重复力为0.909。说明楸树无性系皮孔密度的变异系数和重复力均较大,从中进行选择是比较可靠的。楸树3类皮孔的典型照片见图1。

图1 楸树3类皮孔的典型照片
2.1.2 楸树无性系皮孔的大小
不同无性系间皮孔的大小(即单个皮孔的面积)呈极显著性差异。皮孔最大的无性系为2-2,面积为 11.85 mm2,其次是 9-2,面积为 11.30 mm2,皮孔最小的无性系为011-1,为4.91 mm2,皮孔平均大小为8.50 mm2,变幅为4.91 ~11.97 mm2,遗传变异系数为23.7%。重复力为0.748。皮孔大小分为3类,见表3。第1类:大皮孔(l≥10.10 mm2);第2类:中等皮孔(6.18 mm2≤l≤9.36 mm2);第 3 类:小皮孔(l≤5.94 mm2)。皮孔的大小随树木的生长而变化,树龄越大皮孔的面积也越大,对2年生和5年生楸树无性系的皮孔面积进行年—年相关分析,结果表明皮孔面积呈现出极显著的相关性(R=0.514**)。楸树3类皮孔大小的照片见图2。

图2 楸树3类皮孔大小的照片
2.1.3 皮孔的形状
楸树皮孔的形状近似于椭圆,皮孔突起,但形状差异也较大,对皮孔的长/宽进行聚类分析,将楸树无性系的皮孔划分为3大类,见表4。第1类:狭长椭圆形(sh≥2.85);第2类:椭圆形(1.75≤sh≤2.55);第3 类:近圆形(1.52≤sh≤1.57)。
2.1.4 皮孔性状间的相互关系
通过对29个无性系的皮孔性状与生长指标进行相关分析,结果表明:皮孔密度与皮孔长、皮孔宽、皮孔大小、树高、胸径均呈极显著负相关。即皮孔大的无性系密度较小,皮孔密度小的无性系生长较快,但皮孔的大小与树高、胸径呈极显著正相关(图3和图4),表明皮孔在树木呼吸作用中对树木的生长影响显著,生长好的无性系皮孔大,数量少。总之,皮孔的总面积对树木的生长影响较大,5 cm2面积上的皮孔总面积与树高、胸径呈负相关,这表明皮孔总面积小,减少水分的蒸发,树木生长更快。对无性系的选择提供了依据,即选择优良无性系时,在优先考虑生长指标优良时,尽可能的选择皮孔密度小、皮孔面积小的无性系,达到相对减少水肥供给,实现高生产力的目的,见表5。

表2 楸树无性系皮孔密度识别特征

表3 楸树无性系皮孔大小识别特征

表4 楸树无性系皮孔形状特征
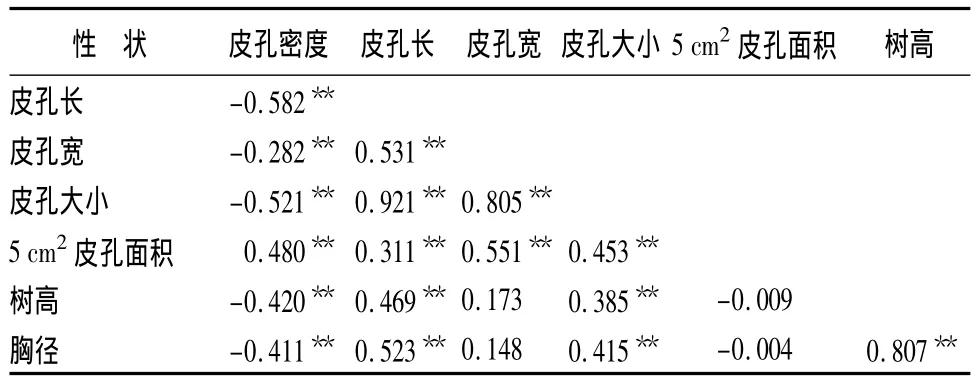
表5 不同无性系皮孔性状与生长指标的相关性
2.2 叶痕性状的变异
2.2.1 楸树无性系叶痕大小的差异
楸树无性系叶间距有很大的差异。无性系间叶痕密度(中部树干1 m段的叶痕数量)、叶痕大小(单个叶痕的面积)呈极显著差异,叶痕密度最大的无性系为011-1,最小的为2-1;叶痕最大的无性系为2-1,最小的无性系为011-1。根据叶痕密度进行聚类分析将叶痕分为3类,见表6:密集型(p≥41.3 个)、较密集型(35.3 个≤p≤39.0 个)、疏散型(p≤34.9个)。叶痕形状呈椭圆形,是3叶轮生。叶痕大小分为3类(表7):大叶痕(lp≥2.92 cm2)、中等叶痕(1.54 cm2≤lp≤2.41 cm2)、小叶痕(lp≤1.50 cm2)。树高、胸径与叶痕面积呈极显著正相关。树高、胸径与叶痕密度呈极显著负相关,表明生长较好的无性系叶痕密度小,但叶痕面积较大。说明叶痕面积大的无性系叶痕密度小,即叶间距大(表8)。
2.2.2 皮孔性状与叶痕性状的相关性
相关分析表明,皮孔密度与叶痕密度呈极显著正相关,与叶痕大小呈显著负相关,说明皮孔密度大的无性系叶痕密度也大。皮孔大小与叶痕密度呈极显著负相关,而与叶痕大小呈极显著正相关,说明皮孔大的无性系叶痕也大。由此可见,楸树无性系的皮孔性状与叶痕性状的变异是一致的。叶痕密度与叶痕大小呈极显著负相关,也与皮孔相一致(表9)。

表6 楸树无性系叶痕密度特征

表7 楸树无性系叶痕大小特征

表8 不同无性系叶痕性状与生长指标的相关性

表9 不同无性系皮孔性状与叶痕性状的相关性
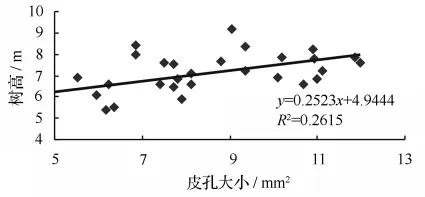
图3 皮孔大小与树高的相关性

图4 皮孔大小与胸径的相关性
2.3 树皮开裂的变异
楸树不同无性系树皮光滑程度不同,有的无性系树皮开裂,有的表面光滑,树皮性状分为两种:开裂型、光滑型。其中树皮开裂的无性系为008-1、002-1、080、线灰4 个;树皮光滑的无性系为:2-2、9-2、015-1、1-3、2-7、2-8、4002、2-1、004-1、4001、9-1、1-4、011-1、6523、灰3、大叶金丝、2-6、1-2、013-1、1-1、小叶金丝、洛灰、光叶楸、038、011-1,25 个无性系(图5)。008-1、线灰呈斑块状开裂,板块大小不同。080、002-1树皮纵向开裂,整个树干全开裂。002-1的树干开裂最严重,开裂程度见图6。从以上分析可以看出,皮孔密度对树干的开裂影响较大,皮孔密度大的无性系树干开裂,无性系002-1与080的密度大树干都开裂。
2.4 楸树无性系各性状综合的聚类分析
楸树无性系表型识别可以从皮孔大小、皮孔密度、叶痕大小、叶痕密度及树皮开裂等性状进行识别,文中分别对各性状的识别进行了研究,对其无性系的特点进行了分类,由于无性系的识别难度较大,为了简单识别每个无性系,对各性状进行综合的聚类分析,将无性系划分为6个大的类群,见图7。
3 结论与讨论
皮孔密度和大小特征用作鉴定植物亲缘关系及抗性,并对楸树优良无性系的早期选择有借鉴意义。在植物遗传育种及资源研究等领域,皮孔性状受到越来越多学者的关注。然而,皮孔特征能否成为鉴定不同无性系亲缘关系指标,研究者尚有争议[12-20]。对29个楸树无性系从皮孔、叶痕、树皮 3个方面进行了研究,对密度及大小进行了分类,皮孔、叶痕密度、大小均分为3类:密度(密集型、较密集型、疏散型);大小(大、中、小)。树皮分类:开裂型、光滑型。楸树皮孔的形状近似于椭圆,皮孔突起,但形状差异也较大,楸树无性系的皮孔划分为3大类:狭长椭圆形、椭圆形、近圆形。
不同无性系间皮孔密度、大小均有极显著性差异,皮孔密度平均为2.836个/cm2,变幅为1.80~5.24个/cm2,遗传变异系数为28.3%。重复力为0.909。皮孔平均大小为 8.50 mm2,变幅为 4.91 ~1 1.97mm2,遗传变异系数为23.7% 。重复力为0.748。楸树无性系皮孔密度、皮孔大小的变异系数和重复力均较大,受遗传因素较大,对于初步鉴定楸树有一定的帮助。皮孔的大小随树木的生长而变化,树龄越大皮孔的面积也越大,对2年生和5年生楸树无性系的皮孔面积进行年—年相关分析表明皮孔面积呈现出极显著的相关性(R=0.514**)。皮孔密度对树干的开裂影响较大,皮孔密度大的无性系树干开裂,例如:无性系 002-1、7080、008-1、线灰的密度大树干都开裂。
无性系间叶痕密度、叶痕大小呈极显著差异。树高、胸径与叶痕大小呈极显著正相关。树高、胸径与叶痕密度呈极显著负相关,表明生长较好的无性系叶痕密度小,但叶痕面积较大。说明叶痕面积大的无性系叶痕密度小,即叶间距大。相关分析表明,皮孔密度与叶痕密度呈极显著正相关,与叶痕大小呈显著负相关,即皮孔密度大的无性系叶痕密度也大。皮孔大小与叶痕密度呈极显著负相关,而与叶痕大小呈极显著正相关,说明皮孔大的无性系叶痕也大。由此可见,楸树无性系的皮孔性状与叶痕性状的变异是一致的。叶痕密度与叶痕大小呈极显著负相关,也与皮孔相一致。

图5 楸树树皮光滑照片

图6 楸树树皮开裂照片

图7 楸树无性系各性状综合聚类
楸树无性系皮孔、叶痕、树皮3个表型性状变异较大,对每个无性系进行详细的区分难度较大,为了简单识别每个无性系,对各性状进行综合的聚类分析,将无性系划分为6个大的类群。3个表型特征能够简单的识别无性系。第1:皮孔特征识别。例如:皮孔密度最大的无性系为002-1,其次是7080。皮孔密度最小的无性系为2-1,其次是015-1。皮孔最大的无性系为2-2,其次是9-2;皮孔最小的无性系为011-1,其次是线灰。第2:叶痕特征识别。例如:叶痕密度最大的无性系为011-1,其次是线灰。叶痕密度最小的无性系为2-1,其次是9-2。叶痕最大的无性系为2-1,其次是4002;叶痕最小的无性系为011-1,其次是小叶金丝。第3:树皮特征识别。例如:树皮开裂的无性系为 002-1、008-1、7080、线灰。树皮光滑的无性系为其它25个无性系。
相关分析表明,皮孔密度与皮孔长、皮孔宽、皮孔大小、树高、胸径均呈极显著负相关。即皮孔大的无性系密度较小,皮孔密度小的无性系生长较快,但皮孔的大小与树高、胸径呈极显著正相关,表明皮孔在树木呼吸作用中对树木的生长影响显著,生长好的无性系皮孔大,数量少。皮孔的形状、大小、分布以及皮孔指标等还直接影响植物对水肥的吸收。因而皮孔的总面积对树木的生长影响较大,5 cm2面积上的皮孔总面积与树高、胸径呈负相关。表明皮孔总面积小,减少水分的蒸发,树木生长更快。皮孔是病菌侵入的主要途径和潜伏的主要部位,皮孔密度及大小与病害发生有密切关系,皮孔总面积大,树皮容易开裂,病菌容易入侵,即选择优良无性系时,在优先考虑生长指标优良时,尽可能的选择皮孔密度小、皮孔面积小的无性系,达到相对减少水肥供给及病害的发生。
[1]姚兆华,郝丽珍,王萍,等.沙芥属植物叶片的气孔特征研究[J].植物研究,2007,27(2):199-203.
[2]姜继芳.YG砧木的生物学特性及对红富士苹果抗轮纹病特性的研究[D].济南:山东农业大学,2009.
[3]Hetherington A M,Woodward F I.The role of stomata in sensing and driving environmental change[J].Nature,2003,424:901 -908.
[4]陈策.苹果果实轮纹病研究进展[J].植物病理学报,1999,29(3):193-198.
[5]陈之端,张志耘.桦木科植物叶表皮的研究[J].植物分类学报,1991,29(2):156-163.
[6]庄东红,宋娟娟.木槿属植物染色体倍性与花粉粒、叶片气孔器性状的关系[J].热带亚热带植物学报,2005,13(1):49-52.
[7]李润唐,张映南,田大伦.柑橘类植物叶片的气孔研究[J].果树学报,2004,21(5):419-424.
[8]郭从俭,张新胜,张万钦.气孔性状对楸树生长及早期选择的影响[J].河南农业大学学报,1996,30(1):65-71.
[9]吴丁,王红,卢金梅,等.中国梅花草属植物的叶表皮特征及其系统学意义[J].植物分类学报,2005,43(3):210-224.
[10]李广旭,沈永波,高艳敏,等.皮孔组织结构及密度与苹果枝干粗皮病发生的关系[J].果树学报,2004,21(4):350-353.
[11]李志英,梁艳荣,胡晓红.梨不同系统叶气孔的密度、大小与起源地气象因子的关系[J].内蒙古农业科技,1994(5):31-32.
[12]张纪英,杨风云.几种落叶果树叶片气孔性状观察[J].邯郸农业高等专科学校学报,2001,18(4):14-15.
[13]杨传友,史金玉,杜欣阁,等.苹果叶片气孔的研究[J].山东农业大学学报,1998,29(1):8-14.
[14]魏岩,周桂玲.十字花科四属植物叶片的表皮特征[J].云南植物研究,1998,20(4):407-412.
[15]曹五七,李逊,谢林,等.杂种棉花叶片气孔形态及数量的扫描电镜观察[J].四川农业大学学报,1995,13(2):158-160.
[16]伊稍·K.种子植物解剖学[M].2版.李正理,译.上海:上海科学技术出版社,1982.
[17]张延龙,牛立新.中国葡萄属植物叶片气孔特征的研究[J].植物研究,1997,17(3):315-319.
[18]铁军,金山,李旭娇,等.濒危植物南方红豆杉叶片形态结构及气孔参数[J].东北林业大学学报,2008,36(9):24-27.
[19]李茂松,王春艳,宋吉青,等.小麦进化过程中叶片气孔和光合特征演变趋势[J].生态学报,2008,28(11):5385-5391.
[20]杨惠敏,王根轩.干旱和CO2浓度升高对干旱区春小麦气孔密度及分布的影响[J].植物生态学报,2001,25(3):312-316.
