白桦与山杨叶凋落物候的差异及其生态学意义1)
2012-06-13栾奎志樊绪富王维芳
栾奎志 樊绪富 王维芳
(黑龙江省林业监测规划院,哈尔滨,150080) (辽宁省海城市林业调查规划院) (东北林业大学)
森林凋落物在维持土壤肥力、促进森林生态系统正常的物质生物循环和养分平衡方面起着重要的作用。一方面,森林凋落物是土壤动物、微生物的能量和物质的来源。凋落物作为养分的有效载体,其在养分循环中是连接植物与土壤的“纽带”,在维持土壤肥力、促进生态系统正常物质循环和养分平衡方面起着重要的作用[1]。另一方面,叶凋落的先后顺序也可能深刻地影响生态系统的物质循环。研究表明,落叶物候还影响树木的养分回收效率[2]。李荣华等[3]对中国南亚热带森林凋落物的分解速率进行了研究,发现不同布设时间的凋落物,其分解速率存在很大差异,认为凋落物的凋落时间可能影响其分解速率;陈金玲等[4]发现在小兴安岭地区,随着进展演替方向,混合凋落叶的年分解系数(k)随之增大;冠层持续时间同时受展叶时间与落叶时间的共同影响,冠层持续时间对树木全年的固碳量影响很大。因此,叶凋落物候对树木的养分循环和碳固定具有重要意义。
Dixon[5]构建了累积凋落量的动态模型,并将该模型应用于北美多个地区的不同树种,并比较了凋落参数的种间和纬度间变化,发现落叶和半落叶树种的凋落高峰时间和凋落期长度存在显著差异,纬度也对凋落速率有影响。随后,Escudero、Del Arco[6]和 Parker 等[7]用此模型研究了不同树种凋落物候的差异。他们的研究不仅证明了叶凋落物动态模型在温带和北方森林的普遍适用性,而且给定量研究不同树种的叶凋落物候提供了有利的工具。
阔叶红松林是我国东北山区的地带性顶极生态系统,目前其原始类型多数已被各种次生类型所代替,其中白桦林和山杨林或其混交的杨桦林是主要的演替早期类型。关于东北地区森林凋落物产量、组成和动态的研究已有不少报道[8-9]。尽管目前对白桦和山杨的研究历史悠久,研究内容也非常广泛,但涉及到二者凋落物候特征的较少。Jin等[8]和原作强等[9]分别研究了小兴安岭和长白山阔叶红松林凋落物组成和季节动态,但是并没有利用凋落物动态模型详细探讨种间的凋落物候差异。白桦(Betula platyphylla)和山杨(Populus davidiana)作为两个主要先锋树种,具有很多相同的生物学特性和生态对策,比如它们叶片的光合速率较高,均为散孔材,且没有明显的心材特征。但是,二者也有不同之处,比如白桦叶片为厚纸质,而山杨则为革质。本研究比较二者叶凋落物候的异同,为进一步研究其他生物学特性提供科学依据和基础数据。
1 研究地概况
本研究的地点设在东北林业大学帽儿山实验林场,位于黑龙江省东南部、尚志市帽儿山镇境内(北纬45°20'~45°25'、东经 127°30'~127°34')。林区平均海拔高度300 m,坡度在10°~15°。该地区年平均气温2.8℃,年均降水量723.8 mm。该地区的植被属长白植物区系,现有植被是原地带性植被——阔叶红松林屡遭人为干扰(采伐、火烧、开垦等)后演替成的天然次生林,代表着东北东部山区天然次生林的典型生态系统。主要植被类型包括沟谷硬阔林、山杨林、白桦林、杂木林和蒙古栎(Quercus mongolica)林,大部分为中龄林。
2 研究方法
分别在白桦林和山杨林分布地段布设5个1 m×1 m凋落物收集器。4—7月每月收集一次凋落物,8月每半月一次,秋季大约10 d收集一次。将收集的凋落物带回实验室,将白桦和山杨叶挑选出来,分别在85℃烘干至恒质量,用电子天平(0.01 g)称质量。
根据Dixon[5]的累积凋落量模型拟合得到凋落物候参数:

式中:F为累积凋落量;P1为最大凋落量(即总叶凋落量);P2为最大日凋落量出现的时间(方程的拐点);P3为从10%到50%凋落量所经历的天数(由于在P2对称,P3也等于50% ~90%凋落量的时间);t表示时间。
图1为累积凋落量模型示意图。该模型实际上包含3个具有生态学意义的参数:凋落的绝对量(P1)、凋落高峰时间(P2,衡量凋落的早晚)和凋落期长度(P3的2倍,即10% ~90%凋落量经历的天数)。由于白桦和山杨的总凋落量不同(分别为213和184 g·m-2),为了比较它们的凋落物候,把最大凋落量(P1)标准化为1或100%进行比较[6]。模型中除了P2和P3外,还可以推算出凋落开始时间、凋落结束时间以及相对凋落峰值(日凋落量最大值)。
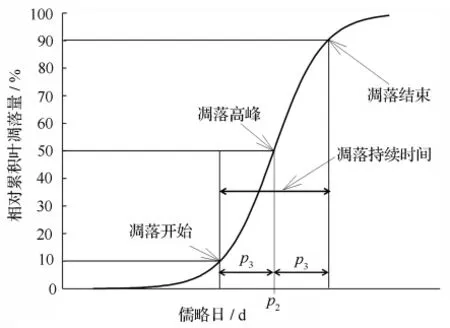
图1 累积凋落量模型示意图
在凋落高峰时从每个凋落物收集器中选取5~10片叶子测定其比叶面积,共5个重复。总叶面积用图像法估计,80℃烘干48 h后用分析天平称质量。叶面积和叶质量为单个叶子的平均面积和质量,比叶面积为单位叶质量的叶面积。C质量分数采用油浴加热重铬酸钾容量法测定,N质量分数采用凯氏定氮法测定。利用SPSS13.0进行数据统计分析。
3 结果与分析
3.1 累积叶凋落动态
累积凋落量模型可以很好地描述两树种的叶凋落动态。每个凋落物收集器收集到的数据的决定系数均大于0.9(P<0.001)。这证明了累积凋落量基本上以凋落高峰时间呈对称分布[2]。尽管白桦和山杨的相对累积凋落量随时间的变化格局均相同,但速率不同(图2)。白桦凋落开始较早,从第240天到第280天凋落速率较高,凋落高峰不明显,凋落期较长;而山杨则表现出完全不同的情况,凋落开始较晚,但迅速结束。但是二者叶凋落期几乎同时结束。

图2 白桦和山杨相对累积叶凋落量的变化
3.2 凋落物候种间差异
方差分析表明(表1):两树种凋落开始时间、凋落高峰时间、凋落期长度和最大相对凋落速率(凋落高峰)差异显著(P<0.001),但二者的凋落结束时间差异不显著(P=0.230>0.05)。具体来说,白桦凋落开始时间(第229天)比山杨的(第270天)早41 d。白桦的平均凋落高峰时间(第256天)比山杨(第276天)早20 d,但二者均晚于北美的垂枝桦(Betula pendula,第 280 天)[5]。白桦凋落结束时间(第282天)比山杨的只早1 d,但统计上不显著(P=0.23)。白桦的凋落期比山杨的长42 d,白桦的相对凋落速率(0.021 d-1)则仅是山杨的(0.099 d-1)五分之一左右(表2)。也就是说,白桦叶凋落时间显著早于山杨,并且白桦叶凋落期较长,其凋落集中程度则明显小于山杨。

表1 白桦和山杨叶凋落物候方差分析结果
原作强等[9]发现,紫椴的叶凋落高峰出现于9月13日—9月26日,且持续时间较长;而蒙古栎、春榆(Ulmus japonica)和色木槭(Acer mono)的叶凋落期则集中于9月27日—10月10日。这4个阔叶树种的凋落高峰介于白桦和山杨之间。在北美,北美红糖槭(Acer rubrum)的凋落高峰出现的时间显著晚于毡毛山核桃(Carya tomentosa)、洋白蜡(Fraxinus pennsylvanica)和北美鹅掌楸(Liriodendron tulipifera)。

表2 白桦和山杨叶凋落物候比较
3.3 凋落叶性状比较
方差分析表明,两树种的叶性状也存在显著差异(表3)。白桦的叶面积、叶质量显著小于山杨的叶面积和叶质量,但白桦比叶面积显著大于山杨。白桦叶N质量分数显著高于山杨,碳氮比值则为白桦显著小于山杨。白桦叶分解速率也明显大于山杨,前者是后者的1.3倍。白桦叶片厚纸质,比叶面积大表明叶片较薄,并且N质量分数高和碳氮比值低,这些都利于白桦叶分解,与白桦分解速率高于山杨相对应。

表3 白桦和山杨凋落叶性状比较
3 结论与讨论
树木的叶凋落物候是树木的重要特征之一。本研究发现,尽管山杨和白桦同为先锋树种,但是它们的叶凋落物候及其相关叶性状具有显著差异。白桦落叶早,落叶期长,落叶分散,而山杨正好相反。这暗示白桦和山杨在叶寿命、冠层叶量、碳吸收和能量储存、养分回收和凋落叶分解等方面可能具有不同的生态对策。
叶凋落物候研究具有重要意义。首先,二者的叶寿命不同。两树种均在5月中上旬展叶[11],但落叶物候相差20~41 d,表明白桦的叶寿命明显比山杨短。实际上,白桦不断地生长新叶,7月份停止叶生长;最先生长的树叶在8月中旬前后就开始凋落,而最后生长的树叶则在10月份才凋落。因此,从整个树木来说,白桦的树叶在生长季内不是一个同生群,而山杨树叶更倾向于一个同生群。其次,白桦在生长季后期(主要是8月下旬和9月份)的冠层叶量(叶面积)会由于凋落明显减少,这直接影响这一阶段的碳吸收。树木一般在8月份停止生长,9月份主要是为度过接下来的冬季和下一年萌芽(或者抵御干扰)积累能量。因此,可以推测白桦在后期的能量积累较少。再次,在叶片衰老之前树木要回收一部分养分以减少其养分损失[12]。叶凋落越早,其氮质量分数越高,养分回收效率越低[2],推测白桦的养分回收效率较低。最后,由于早期凋落物的氮质量分数较高[1],碳氮比较低,因此,早期凋落的树叶从化学性质上看易于分解。而凋落早期水热条件也较好,可以预测白桦早期凋落叶分解较快,这直接影响到白桦凋落叶的分解和养分释放[4]。白桦叶分解速率是山杨的1.3倍也间接证明了这一推测。由于本研究只涉及两个树种,还不能得出凋落早的树种的叶分解比凋落晚的树种快的结论。
本研究利用凋落物动态模型定量研究了白桦和山杨的叶凋落物候及其叶性状,发现两个树种的叶物候及相关对策存在显著差异,为深刻认识两个树种的林学特性提供了基础数据。
[1]王凤友.森林凋落量研究综述[J].生态学进展,1989,6(2):82-89.
[2]Niinemets¨U,Tamm¨U.Species differences in timing of leaf fall and foliage chemistry modify nutrient resorption efficiency in deciduous temperate forest stands[J].Tree physiology,2005,25(8):1001.
[3]李荣华,邓琦,周国逸,等.起始时间对亚热带森林凋落物分解速率的影响[J].植物生态学报,2011,35(7):699-706.
[4]陈金玲,金光泽,赵凤霞.小兴安岭典型阔叶红松林不同演替阶段凋落物分解及养分变化[J].应用生态学报,2010,21(9):2209-2216.
[5]Dixon K.Analysis of seasonal leaf fall in north temperate deciduous forests[J].Oikos,1976,27(2):300-306.
[6]Escudero A,Del Arco J M.Ecological significance of the phenology of leaf abscission[J].Oikos,1987,49(1):11-14.
[7]Parker G G,O’Neill J P,Higman D.Vertical profile and canopy organization in a mixed deciduous forest[J].Plant Ecology,1989,85(1):1-11.
[8]Jin G Z,Zhao F X,Liu L,et al.The production and spatial heterogeneity of litterfall in the mixed broadleaved-Korean pine forest of Xiaoxing’an Mountains,China[J].Journal of Korean Forest Society,2008,97(2):165-170.
[9]原作强,李步杭,白雪娇,等.长白山阔叶红松林凋落物组成及其季节动态[J].应用生态学报,2010,29(9),2171-2178.
[10]李雪峰.长白山森林生态系统碳过程的研究[D].哈尔滨:东北林业大学,2003.
[11]祝宁,江红,金永岩.中国东北天然次生林主要树种的物候研究[J].植物生态学与地植物学学报,1990,14(4):336-349.
[12]孙书存,陈灵芝.东灵山地区辽东栎叶养分的季节动态与回收效率[J].植物生态学报,2001,25(1):76-82.
