凡纳滨对虾养殖水体中浮游植物群落的组成分析
2012-06-08李由明黄翔鹄李晓梅
李由明,黄翔鹄,李晓梅
(1琼州学院生命科学与技术学院,海南三亚572022;2广东海洋大学水产学院,广东湛江524088)
浮游植物作为水体中初级生产力的代表,在物质循环过程中扮演者提供物质能量的角色,同时浮游植物还是水体中溶解氧的主要来源.在对虾养殖过程中,高密度的集约化养殖方式,导致养殖水体微环境的不稳定性.浮游植物群落组成的变化是水体环境变化的一个反应指标,其对养殖过程中营养盐的调控和维护养殖水体中微生态环境的稳定具有重要作用[1,2].近些年,对虾养殖过程中疾病的频发,引起了人们对养殖环境稳定性调控的关注.虾池中的浮游植物对水体中营养盐的吸收,在一定程度上使得养殖水体的水质保持良好,在一定程度上防止了水体富营养化[3].
对凡纳滨对虾养殖水体系统中浮游植物群落的组成与演替变化进行了调查与分析,为进一步利用藻类对养殖水体环境进行生物调控研究提供基础数据.
1 材料和方法
1.1 试验所用虾池
实验选择三个不同类型凡纳滨对虾养殖池塘,选用虾池为铺有聚乙烯塑料薄膜的高位池(一种高密度的集约化对虾养殖模式池塘).池塘底部高于海水的最高潮位线1-6m,完全依靠动力供水,每个池塘面积大小为2.67 ×103m2/个,水深1.60 -1.75m,放养密度为1.50 ×107~2.25 ×107尾/hm2.每口虾池配备 0.75kW的4个叶轮式增氧机和一个潜水式增氧机,水体为循环水.试验期间水温为19~23℃,盐度14~20.
1.2 虾池中浮游植物的测定
1.2.1 样品采集和处理
试验期间采样时间15:00-17:00,每两天采样1次.在每个池塘的上风口、下风口和中间各选择一个采样点.在每一采样点的水表层下30~40cm深度采水样1L,现场加15mL鲁哥氏液固定.
将固定的水样置于室内分液漏斗中静止沉淀,在水样沉淀开始2h后将分液漏斗轻轻旋转,防止藻类附着在分液漏斗内壁,沉淀48h,浓缩至50mL,并加少量的甲醛保存.
1.2.2 样品分析
将水样用左右平移的方式轻轻摇动200次,摇匀后立即用定量吸管从中吸取0.1mL置入0.1mL计数框内,在40倍的物镜下观察计数,每个水样计数两次,取其平均值.具体观察的视野数以样品中浮游植物的多少而定;如果平均每个视野有十几个到几十个,50个视野即可.如果每个平均每个视野有5-6个时,需要100个视野;如果平均每个视野不超过1-2个时,要200个视野.在对样品定量分析的同时,对样品进行定性分析.
2 结果
2.1 藻类的种类组成
通过不同的凡纳滨对虾养殖水体取样分析,水体中藻类的主要实验虾池中的藻类共属于5个门,21个种.其中,硅藻门(Bacillariophyta)有9种,占所有藻类种类数的42.86%,分别为:小环藻(Cyclostella sp.)、直链硅藻(Melosira sp.)、舟形藻(Navicula sp.)、角毛藻(C.mueeleri sp.)、新月拟菱形藻(N.closterium sp.、桥穹藻(Cymbella sp.)、菱形藻(Nitzschia sp.)、四棘藻(Atthetas sp.)、平板藻(Tabellaria sp.);绿藻门(Chlorophyta)有5种,占所有藻类种类数的23.81%,分别为:衣藻(Chlamydomonas sp.)、波吉卵囊藻(Oocystis borgei、)小球藻(Chlorella sp.)、绿球藻(Chlorococcum sp.)、纤维藻(Ankistrodesmus sp.);蓝藻门(Cyanophyta)有5种,占所有藻类种类数的23.81%,平裂藻(Merismopedia sp.)、席藻(Phormidium sp.)、色球藻(Chroococcus sp.)、螺旋藻(Ppirulina sp.)、颤藻(Oscillatoria sp.);甲藻门(Pyrrophyta)有 1 种,为沟环藻(Gymnodinium sp.),占所有藻类种类数的4.76%;隐藻门(Cryptophyta)有1种,为隐藻(Cryptomonas sp.),占所有藻类种类数的4.76%.
2.2 优势种群组成
1号池塘水体中浮游植物优势种为绿藻门的小球藻(Chlorella sp).和波吉卵囊藻(Oocystis borgei);硅藻门的小环藻(Cyclostella sp);2号池塘水体中藻类的优势种为绿藻门的波吉卵囊藻Oocystis borgei),硅藻门的小环藻(Cyclostella sp)和新月拟菱形藻(N.closterium sp),蓝藻门的平裂藻(Merismopedia sp.和席藻(Phormidium sp);3号池水体中藻类的优势种绿藻门的波吉卵囊藻(Oocystis borgei),硅藻门的小环藻(Cyclostella sp)、新月拟菱形藻和角毛藻(C.mueeleri sp),蓝藻门的平裂藻(Merismopedia sp)和席藻(Phormidium sp).(见表5)
虾塘水体中早期的浮游植物优势种群为绿藻门的小球藻和波吉卵藻藻,硅藻门的小环藻、新月拟菱形藻、角毛藻,蓝藻门的平裂藻和席藻.其中,波吉卵囊藻的数量在试验16d至19d达到最大,最大数量为2.35×107个/升,其在藻类总的数量中所占比例最大为98.09%;小球藻的数量在试验早期开始时最大,最大数量为6.87×105个/升,在藻类数量中所占比例最大为26.6%;小环藻数量的在试验早期最大,最大数量为4.79×106个/升,在藻类数量中所占最大比例为59.6%;新月拟菱形藻的数量在13d最大,最大数量为2.82×107个/升,在藻类数量中所占比例最大为85.21%;角毛藻的数量在13d达最大;最大数量为2.80×106个/升,平裂藻的数量在10d达最大,最大为8.11×105个/升,在藻类数量中所占比例最大为5.8%;席藻的数量在10d达最大,最大为1.1×106个/升,在藻类数量中所占比例最大为27.87%(见表2~表4).
2.3 优势种群的数量变化
对虾养殖水体中浮游植物优势种数量变化范围为3~6个.在养殖早期多数优势种类的数量波动较小,到养殖的中后期,优势种类的数量变大,相对早期,数量波动也较大(见图1~图5).

图1 1号池水体中小球藻和小环藻种群数量变化
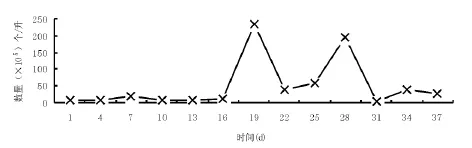
图2 1号池中波吉卵囊藻种群数量变化

图3 2号池水体中浮游植物优势种群的数量变化

图4 3号池浮游植物优势种群的数量变化
3 讨论与分析
3.1 浮游植物群落的变化
在虾苗投放到虾池前,水体中浮游植物群落自然的演替,水体环境相对稳定,使得藻相具有相对高的稳定性[5].虾苗投放到虾池中后,对虾池中水环境的理化因子以及水体中食浮游植物的浮游动物的种群产生了影响,从而影响了浮游植物群落的种类和数量。虾体幼体对浮游植物群落的影响主要是通过其对水体中浮游动物种群的调控,从而间接的调控水体中浮游植物的群落[6];同时,对虾幼体还滤食一些浮游植物,进而对浮游植物群落变化产生直接的影响.从观察的不同时间藻类的组成和数量的变化可知,1号池和3号池的藻类种类数波动较大,说明在虾苗投放到水体中后的一段时间内,水体中的藻相变的不稳定.而2号池由于在虾苗投放前,水体中藻类种类数较多,在虾苗投放后,其藻类种类数波动较1号池和3号池小,说明藻相较另两个池稳定.所以,从这三个虾池虾苗投放前后水体中浮游植物群落的变化可知,养殖水体中浮游植物群落早期的结构特征对保持养殖后期水体中藻相的变化具有重要的影响作用.
3.3 浮游植物优势种群的变化
在优势种群中,小球藻和小环藻的种群变化趋势是基本是一致的,并且这两种作为优势种群的藻类比较稳定.一方面可能是在养殖早期水质变化不大,水环境比较稳定.另一方可能是这两种由藻类的生理特征决定的.查广才等[4]对凡纳对虾淡化养殖过程中微型浮游生物群落及多样性进行研究后,也是发现在养殖早期小环藻是养殖水体中的优势种之一.
波吉卵囊藻在实验的前半阶段,即在虾苗投放到高位池一个月左右的时间内,其种群稳定(图2).波吉卵囊藻种群在实验阶段的后半时期,即虾苗放到高位池一个月到两个月之间的时间内,其种群变动较大,主要原因是水体中氨氮的浓度数值也是在此阶段内呈增加的趋势(见另文,待发表),故波吉卵囊藻种群变化与水体中氨氮的浓度有关.
从虾体水体中浮游植物优势种群的变化曲线知(见图1至图4),在对虾养殖过程中,虾塘水体中浮游植物的主要优势种群为绿藻门的波吉卵囊藻和小球藻以及硅藻门的小环藻.这与黄翔鹄等[7]对虾塘水体中浮游植物种群的研究结果一致.从虾塘水体中浮游植物的优势种组成可以得出结论:在虾塘水体中,浮游植物主要为绿藻门、硅藻门和蓝藻门中的一些种类;群落特征为优势种群单一、突出.
[1]孙耀,宋云利.虾塘养殖水环境中氮磷营养盐的存在特征与行为[J].水产学报,1998,6(22):117-123.
[2]孙耀,李锋,李键等.虾塘水体中浮游植物群落特征及其与营养状况的关系[J].海洋水产研究,1998,19(2):45-51.
[3]卢静,李德尚.对虾池的放养密度对浮游生物群落的影响[J].水产学报,2000,24(3):240-246.
[4]查广才,周昌清,黄建荣等.凡纳淡化养殖虾池微型浮游生物群落及多样性[J].生态学报,2004,8(24):1748-1755.
[5]郭皓,于占国.虾池浮游植物群落特征及其与虾病的关系[J].海洋科学1996,20(1):39-44.
[6]胡鸿均,李荛英,魏印心等.中国淡水藻类[M].上海:上海科学技术出版社,1980:10-478.
[7]黄翔鹄,王庆恒.对虾高位池优势浮游植物种群与成因研究[J].热带海洋学报,2002,10(21):36-44.
