东祁连山高寒草地土壤3种高山被孢霉生物学特性的测定
2012-05-13高晓星陈秀蓉
高晓星,陈秀蓉
(甘肃农业大学 草业学院/草业生态系统教育部重点实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
高山被孢霉是一种丝状的腐生菌,含有丰富的多糖和蛋白质。广泛分布于土壤内、牲畜的粪便中及其他腐败的有机物质上[1]。1987年Totani首次发现高山被孢霉,隶属于接合真菌亚门。作为自然界的拓荒者,可以直接发酵生产糖化酶和蛋白酶类等,同时,还可以发酵生产(GLA)等有机酸[2]。根据医学研究相关报道,GLA具有诸多药用价值和营养价值,对糖尿病、心脏病、高血压、高血脂、中风、过敏性湿疹等疾病都具有明显的疗效、可促进被酒精损伤肝功能的恢复,对免疫系统疾病具有调节作用[3,4]。另外,由于高山被孢霉的生长繁殖速度快、资源丰富、油脂含量高、生长周期短、碳源利用谱广和不受气候限制等特点,这就有利于实现大规模生产,这一工业生产研究就是要加快微生物油脂发酵技术创新和产业化进程,可为我国未来生物柴油产业化和油脂化工行业的健康发展提供保障。随着现代生物技术的发展使产油微生物的研究技术在不断趋向成熟。1988年Shimizu等[5]认为高山被孢霉是可以进行商业化生产的一个潜在资源,同时,高山被孢霉也被认为是产生多价不饱和脂肪酸的优良微生物[6,7],国内对高寒草地土壤真菌的研究刚刚起步,而高山被孢霉是东祁连山高寒草地土壤真菌中的优势种群[8]。另外,高山被孢霉用于产业化生产所要求的指标不仅要合成高水平的不饱和脂肪酸酯,也要有较高的生物量,要达到此目的,适宜的营养成份及配比对提高孢子萌发率、增加菌丝分枝和强壮菌丝体都是十分重要[9]。因此,研究以3种高山被孢霉为对象,单因素对不同碳氮源、温度、pH、湿度及光照强度等培养条件进行摸索,以期找到高山被孢霉菌丝生长最佳的培养条件,从而为高山被孢霉的进一步研究提供理论依据。
1 材料和方法
1.1 供试菌株
供试菌株M.ap 27,M.ap 60和M.ap 9是由课题组从东祁连山高寒草地土壤中分离获得,由甘肃农业大学草业学院微生物实验室保存。
1.2 供试培养基
PDA培养基由马铃薯200g、琼脂8g、葡萄糖20g、H2O 1 000mL,用于3种高山被孢霉菌种活化;察氏培养基NaNO33g、K2HPO41g、MgSO4·7H2O 0.5g、KCI 0.5g、FeSO40.01g、30g、琼脂20g、H2O 1 000mL,用于供试菌株生长温度和pH的筛选测定;基础培养基为FeSO40.01g、KCI 0.5g、K2HPO41g、琼脂8g、H2O 1 000mL,用于高山被孢霉利用碳、氮源的对照;供试碳源培养基是在基础培养基中,加入0.2%NaNO3,再分别加入3% 葡萄糖、麦芽糖、蔗糖、淀粉、乙酸钠等碳源,各处理重复3次;供试氮源培养基也是在基础培养基中,加入3%葡萄糖,再分别加入0.2%KNO3、尿素、氯化铵、蛋白胨、硝酸铵等氮源,各处理重复3次。
1.3 试验方法
1.3.1 pH值对菌落生长的测定 从实验室挑选出3株高山被孢霉接种到PDA培养基上活化培养,用直径0.6cm的打孔器切取菌饼,分别接种于察氏培养基,制备好的PDA培养基利用酸度计,通过0.1mol/L NaOH溶液和0.1mol/L HCl溶液将培养基浓度调节为4、5、6、7、8、9不同的6个梯度,再将活化好的供试菌株分别点接在培养基上,各处理重复3次。25℃恒温培养7d,利用十字交叉法测量并记录菌落直径。连续测量5次,计算出菌落的平均生长速率。所得数据用DPS软件进行方差分析。
1.3.2 光照强度对菌落生长的测定 将活化好的供试菌株接到察氏培养基之后,置于可控光照培养箱内,分别将光照强度设置为200lx、400lx、600lx、800lx,以没有光照的处理为对照。培养方法同1.3.1所述。
1.3.3 湿度对菌落生长的测定 将3种高山被孢霉接种到察氏培养基中,利用浓硫酸将湿度调节为10%、25%、35%、50%、65%、75%、90%(表1)[10],培养方法同1.3.1所述。

表1 硫酸浓度与相对湿度的关系Table 1 The relationship of sulfuric acid concentration and relative humidity
1.3.4 3种高山被孢霉对多种碳源、氮源利用能力的测定 将待测菌株点接到供试碳源培养基或氮源培养基上,其中,供试碳源有葡萄糖、麦芽糖、蔗糖、淀粉、乙酸钠;供试氮源有KNO3、尿素、氯化铵、蛋白胨、硝酸铵。各处理重复3次,然后置于25℃恒温培养箱内培养,以基础培养基作为对照(ck)。培养方法同1.3.1所述。
2 结果与分析
2.1 pH对菌丝生长的影响
pH对菌丝生长的影响结果(表2)。不同pH处理下3种高山被孢霉都能生长,其中,M.ap 27在中性偏酸性(pH=5~7)的培养基上菌丝生长速率都大于0.9cm·d-1,并与在碱性培养基(pH 8~9)具有显著差异;M.ap 60和M.ap 9的最适宜生长pH为7。另外,在pH 为7时,M.ap 60的生长速率(1.12cm/d)要大于 M.ap 27(0.96cm/d)和 M.ap 9(0.60cm/d)的生长速率。可见,3种高山被孢霉对酸碱的适应范围较广,而M.ap 60对的土壤酸碱有更好的适应性。

表2 pH对3种高山被孢霉菌丝生长的影响Table 2 Effects of pH on hypha growth of three Mortierella alpina strains
2.2 光照强度对菌丝生长的影响
光照强度对菌丝生长的影响(表3),在弱光强(200~600lx)的处理下,M.ap 27的生长速率与光照强度成正比,光照强度越大菌丝生长速率越大,其中,光强为600lx时,M.ap 27的生长速率最快,达到0.71 cm/d,与对照之间(0.57cm/d)存在显著差异;M.ap 60的生长速率在600lx的光照下达到最大值,为0.67 cm/d,M.ap 9在400lx的光强下生长速率为0.74 cm/d,与其他处理之间存在显著差异。
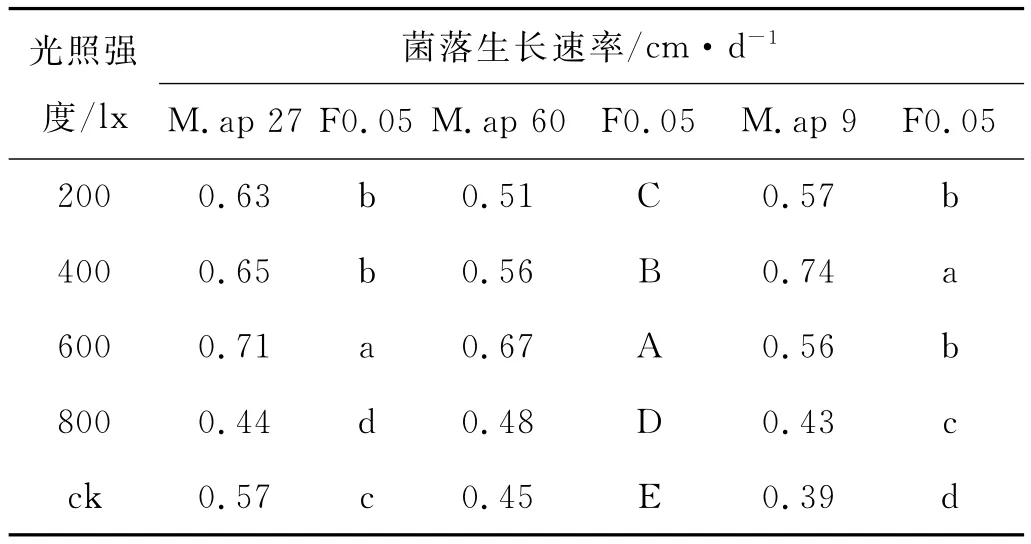
表3 光照强度对3种高山被孢霉菌丝生长的影响Table 3 Effects of Light intensity on hypha growth of three Mortierella alpina strains
2.3 湿度对菌丝生长的影响
湿度对菌丝生长的影响结果(表4),在湿度小于75%时,M.ap 27的生长速率随着湿度的增加呈增长趋势,而湿度达到90%时就会抑制菌丝的生长速率。当湿度为75%时,3种高山被孢霉的菌丝生长速率达到最大值,可见湿度过低或过高都会抑制菌丝的平均生长速率。

表4 湿度对3种高山被孢霉菌丝生长的影响Table 4 Effects of humidity on hypha growth of three Mortierella alpina strains
2.4 菌丝对多种碳源、氮源的利用能力
3种高山被孢霉在不同的碳源培养基上具有显著差异。其中,M.ap 27在淀粉和蔗糖培养基上菌丝生长速率最快,分别为1.06cm/d和0.99cm/d,与对照差异显著,最适碳源为淀粉,麦芽糖和葡萄糖对菌落生长的影响与对照(0.77cm/d)差异不显著,而在乙酸钠的处理下,菌丝生长速率为0.35cm/d,对菌丝生长有显著的抑制作用。M.ap 60在蔗糖与淀粉培养基上菌丝生长速率在0.92~1.0cm/d,与对照之间存在显著差异,而麦芽糖、葡萄糖培养基与ck之间没有差异,乙酸钠对其生长有抑制作用;M.ap 9在葡萄糖培养基上生长速率最快,为0.61cm/d,可见,M.ap 9的最适碳源为葡萄糖,除麦芽糖对其生长有促进作用外,其他碳源对菌丝生长的影响与对照都没有显著差异。因此,3种高山被孢霉在各自最适的培养条件下,M.ap 27的生长速率最快(表5)。

表5 不同碳源对3种高山被孢霉菌丝生长的影响Table 5 Effects of C-source on hypha growth of three Mortierella alpine strains
3种高山被孢霉的最适宜氮源都为蛋白胨,对于M.ap 27而言,其他氮源也都有促进菌丝生长的作用,并且与对照之间具有显著差异;氯化铵和硝酸铵对M.ap 60的生长仍然有促生作用,而硝酸钾和尿素对其生长的影响与对照之间没有显著差异;硝酸钾对M.ap 9的菌丝生长有促生作用,而氯化铵对其有抑制作用,M.ap 9对各种氮源的利用能力与对照之间差异都不显著(表6)。
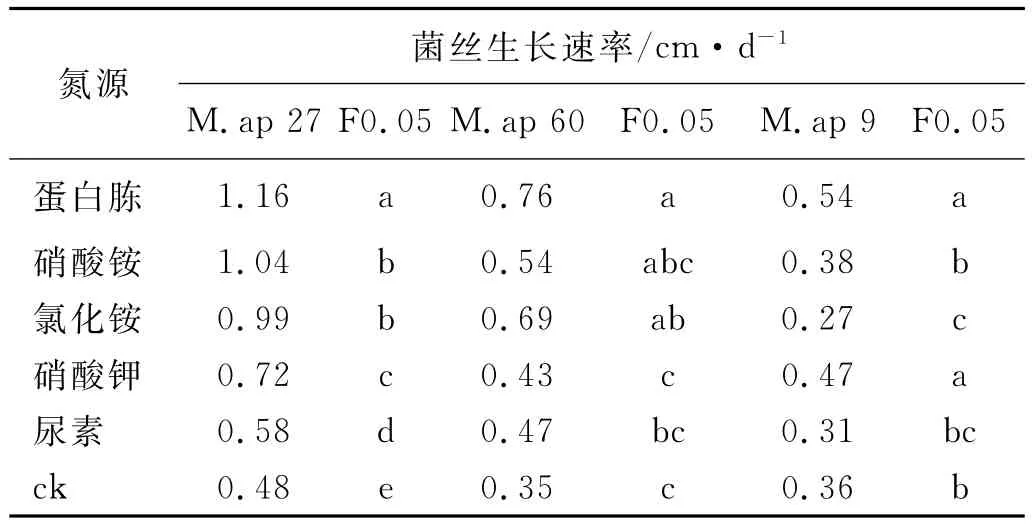
表6 不同氮源对3种高山被孢霉菌丝生长的影响Table 6 Effects of N-source on hyphae growth of three Mortierella alpina strains
3 讨论与结论
试验从pH、光照强度、湿度及碳氮源等方面对3种高山被孢霉的生物学特性进行测定,结果表明不同处理下3种高山被孢霉的生物学特性具有显著差异。3菌株在pH 4~9范围内均能生长,最适宜pH为7;M.ap 27的最适碳源为淀粉,M.ap 60的最适碳源为蔗糖,M.ap 9的最适碳源为葡萄糖;M.ap 27,M.ap 60和M.ap 9的最适氮源均为蛋白胨。3种高山被孢霉在基础培养基上都能生长,证说明有较强的适应贫瘠营养环境的能力。供试菌株在6种碳氮源培养基上均可生长,表明供试菌对碳源利用较为广泛,不仅能利用单糖等速效碳源,而且还能利用淀粉等长效碳源,说明其在加速凋落物的降解过程中发挥着重要的作用[11]。另外,不同种的高山被孢霉对同一种碳源或氮源的利用能力不同,这说明不同种高山被孢霉由于生理特性有差异,对碳或氮源的要求不同[12]。由于在初试条件下3种高山被孢霉能够在0~35℃下存活,故将温度设定为25℃,但实验结果表明3供试菌株的菌丝生长速率之间存在很大差异,可能与供试菌的最适温度有关,而这也是本试验的没有考虑到的不足之处。因此,对于高寒草地土壤中高山被孢霉的研究还需要从更多方面的生物学特性及功能进行试验,为产脂微生物的工业发酵产业及医学方面的应用提供依据。
[1] 诸葛健,李华钟.微生物学[M].北京:科学出版社,21世纪高等院校教材-生物工程系列,2004(9):102-105.
[2] 姜成林,徐丽华.微生物资源开发利用[M].北京:中国轻工业出版社,2001,85-86.
[3] 朱国胜,刘作易,雷邦星,等.被孢霉-亚麻酸高产菌株选育[J].菌物学报,2005(1):85-92.
[4] 周晓兰,李斌,施碧红,等.23种生化因子对被孢霉Mor-tierellal sabelinaF-898生长代谢的影响[J].福建师范大学学报(自然科学版),2001,17(3):92-95,120.
[5] Shimizhu S,Kawashia H,Akimoto K,et al.Production of Ecosapentaenoic acid by Mortierella fungi[J].JAm Oil ChemSoc,1988,65(1):445-459.
[6] 黄建忠,施巧琴,周晓兰,等.高产脂微生物深黄被孢霉M018变株的选育及其油脂合成[J].福建师范大学学报(自然科学版),1998,8(4):233-237.
[7] 铃木修.C2亚麻酸的微生物生产[J].发酵与工业,1983,43(11):1024-1031.
[8] Pointing S B,Buswell J A,Jones E B,et al.Extracellular cellulolytic enzyme profiles of five lignicolous mangrove fungi[J].Mycological Research,1999,103:696-700.
[9] 林琳,杨梅,黄谚谚,等.提高灰黄霉素产生菌Penicillium patulumF2208孢子萌发率的生化因子[J].福建师范大学学报(自然科学版),1996,12(2):88-92.
[10] 方中达.植病研究法(第三版)[M].北京:中国农业出版社,1998:110,343-345.
[11] 王国荣.东祁连山高寒灌丛草地土壤微生物的季节动态和生态分布[D].甘肃农业大学硕士论文,2005.
[12] Raghukumar C,Raghukumar S,Chinnaraj A,et al.Laccase and other lignocellulose modifying enzymes of marine fungi isolated from the coast of India[J].Soil Biochem,1994,37:515-523.
