基于STELLA模型的天敌-稻纵卷叶螟系统的情景分析*
2012-05-09章家恩骆世明李逸勉
秦 钟,章家恩,骆世明,张 锦,李逸勉
(华南农业大学农学院生态学系∥华南农业大学农业部生态农业重点开放实验室∥广东省高等学校农业生态与农村环境重点实验室,广东 广州 510642)
害虫种群数量动态的研究是种群生态学研究的重点内容之一。20世纪60年代以来,随着害虫防治技术和研究方法的不断发展,将系统科学理论和方法引入到种群数量动态及控制的研究中,从而实现系统的优化控制为害虫的防治研究提供了新途径。Watt[1-2]、Morris[3]等是率先应用系统分析方法研究昆虫种群动态的科学家,Huffaker[4-5]在其主持的有害生物协调管理计划中也特别强调应用系统处理方法研究各种害虫及病害的重要性。美国、加拿大、荷兰等国家应用系统科学方法和模拟技术先后对苜蓿、大豆、棉花、柑桔等作物系统的害虫综合治理进行了颇具成效的研究。国内学者如丁岩钦[6]、李典谟[7]、尹长民[8]等在20世纪80年代对系统分析方法应用于害虫种群系统控制途径、方法等进行了探讨和实践。庞雄飞[9-10]、徐汝梅等[11]从系统科学的观点出发,提出以昆虫种群为对象系统,将作用于种群的各种因子经信息处理成为对种群发育过程产生影响的控制信号,通过信息的作用及反馈,研究种群数量控制问题,并运用生命表分析、网络模型、矩阵模型、昆虫种群系统的状态空间表达式、多种群相互作用共存系统的信息处理等方法,对害虫种群系统控制进行了深入研究。
系统动力学(System Dynamics,简称SD)是系统科学的一个重要分支,也是系统科学理论与计算机仿真紧密结合、研究系统反馈结构与行为的一门科学,其主要特点就在于从系统结构入手,将研究对象划分为若干子系统,建立各个子系统之间的因果关系网络,将系统与环境、系统内部各子系统之间相互作用的复杂关系通过一系列微分方程和函数关系加以表述,从而实现对系统结构、功能乃至发展趋势的定量模拟和预测[12-14]。自20世纪50年代后期诞生以来,这一方法的理论与应用研究已遍及各类系统,涉及多种学科和领域,特别适合于分析解决社会、经济、生态和生物等一类非线性复杂大系统的问题[15],但目前这种方法在国内昆虫种群动态方面的研究并不多见。
稻纵卷叶螟是广泛分布于我国稻区,尤以南方稻区密度为大的一类害虫,近年来,随气候、耕作制度、栽培条件的变化,呈现虫量大、世代重叠严重,危害现象明显等特点[16]。从害虫种群系统控制的角度出发,以稻田天敌─稻纵卷叶螟种群系统为研究对象,分析该系统结构、功能与动态行为的内在关系,利用系统动力学的相关理论和方法构建仿真模型,通过改变影响系统行为特性的控制参数,对不同的稻纵卷叶螟种群控制方案进行仿真,可以获取更丰富的信息,进而探寻害虫控制的最佳途径[17]。本研究即以此为出发点,借助系统动力学的分析工具─ STELLA9.0(Strongly-Typed,Lisp-like Language,简称STELLA)对天敌─稻纵卷叶螟种群系统进行模拟,研究结果不仅为稻纵卷叶螟种群的数量预测、优化控制及稻田生态系统的田间管理提供科学依据,同时也将是系统科学在害虫综合管理中应用的深入实践和有益补充。
1 研究方法与步骤
1.1 构建概念模型
稻纵卷叶螟系迁飞性害虫,在广东省中部稻区,年发生7~8个世代,以幼虫在禾本科杂草中越冬,每年春季越冬幼虫化蛹羽化,以后约每隔1个月左右发生1代。第1代稻纵卷叶螟为田间存留的零星成虫,发生在3-4月份。第2代为迁入代,发生在4月初至5月中旬,其幼虫为害分蘖期稻株。第3代是为害的主要世代,即盛发代,发生在5月下旬至6月中旬。第4代发生在6月下旬到7月下旬,为继发代,常为害迟插迟熟品种和晚稻秧苗。晚稻稻纵卷叶螟虫源来自8月中旬至9月中旬第五代成虫的大量迁入,第六代是晚稻的主要为害世代,第7,8代虫口密度很低,基本上不造成危害[18]。考虑到稻纵卷叶螟在早稻和晚稻生长期有着相似的发生过程,即迁入代-盛发代-继发代3个主要发生世代,本研究仅以早稻田间发生情况进行模拟。
在稻纵卷叶螟发育过程中,不同的发育阶段具有明显的形态特征,对作用于种群的各因子可能发生不同的响应。为此,将稻纵卷叶螟种群在早稻生育期间的3个世代作为研究阶段,各世代随着时间的推移以一定的数量转移关系作为纽带依次联结在一起,影响稻纵卷叶螟种群发育状态的各个因子分别作用于状态间的转移关系,以此为基础开始种群生命系统的模拟过程。另一方面,由于稻纵卷叶螟的发生与危害程度与天敌有着极为密切的关系,为便于研究,可将稻纵卷叶螟的寄生性天敌和捕食性天敌作为天敌系统,分析天敌系统与不同世代的稻纵卷叶螟种群系统之间的寄生与被寄生、捕食与被捕食关系,不考虑天敌、稻纵卷叶螟的种内和种间竞争,也不考虑其它因素如气候的影响,由此建立的概念模型如图1所示。

图1 稻纵卷叶螟-天敌系统的概念模型
根据稻纵卷叶螟在早稻生育期内的世代发育进程,将第2,3,4代稻纵卷叶螟及天敌系统均视为“库(stock)”。从稻纵卷叶螟成虫迁入,经卵期-幼虫期-蛹期-下一代成虫,直至第4代成虫迁出,前一虫态向后一虫态的依次转化可视为“流(flow)”,虫态转化的密度由前一虫态的密度、存活率(或每雌产卵量)决定。为了便于研究,将稻纵卷叶螟成虫向下一代成虫转化的中间过程用STELLA软件中的“输送带(conveyor)”来表示,成虫在“输送带”停留的时间即为稻纵卷叶螟完成一个世代所需的天数。影响“库”、“流”状态或变化的参数 (或常数)为“转换器(converter)”,如文中天敌对稻纵卷叶螟的攻击系数可作为稻纵卷叶螟种群密度的主要控制变量。根据稻纵卷叶螟各世代之间及与天敌系统之间的相互联系、相互制约关系及其动态变化特征,利用STELLA软件构建相应的模型(图2)。
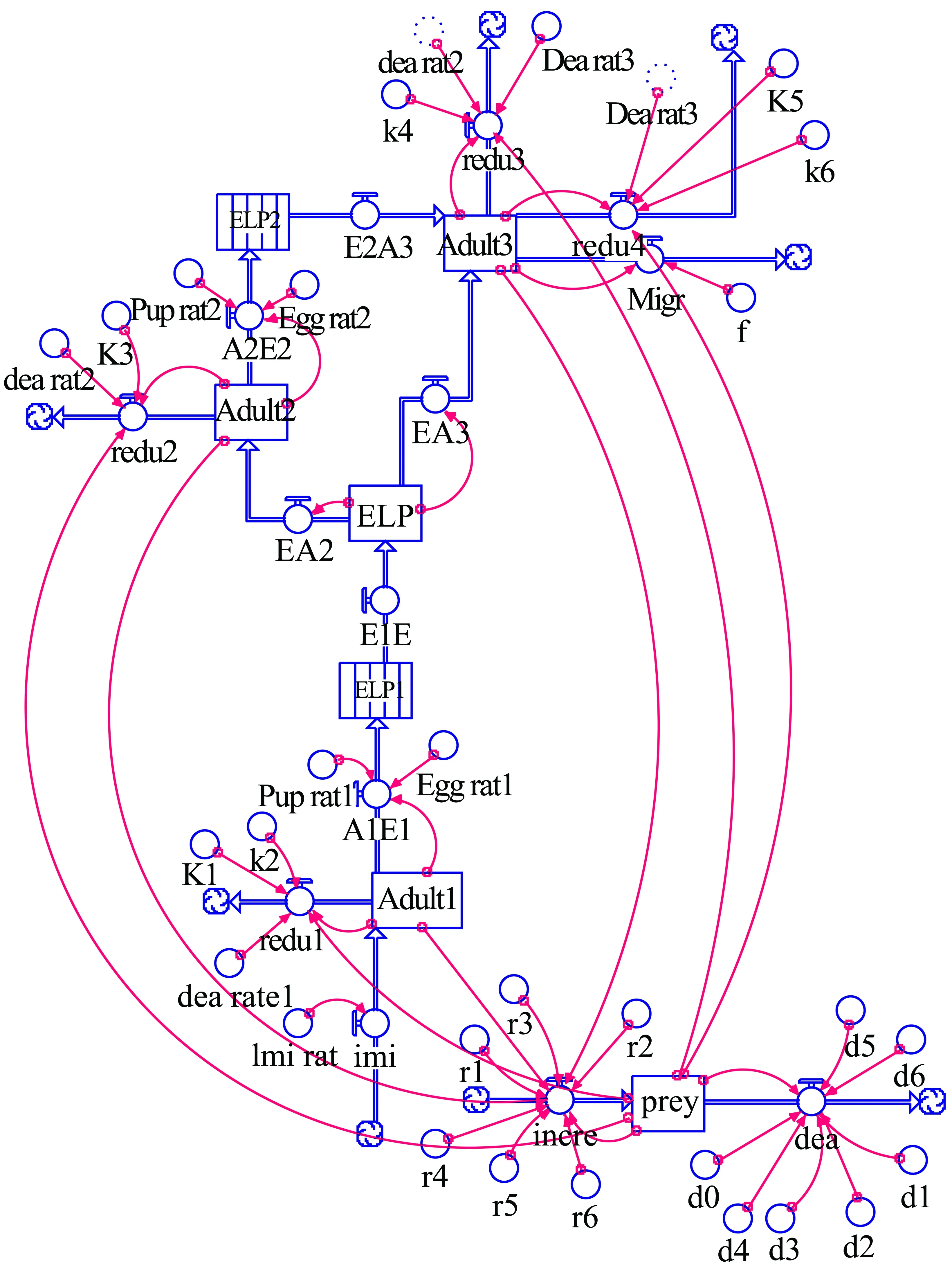
图2 稻纵卷叶螟-天敌系统的STELLA模型
1.2 模型的主要过程及方程
假设水稻的生育期为100 d,稻纵卷叶螟从某时刻t1开始迁入,迁入历期为t1≤t 式中r表示稻纵卷叶螟被天敌成功捕获并经消化、吸收后,对天敌自身增长的贡献系数(以下简称贡献系数),d表示天敌在某时刻的自然死亡率。为了研究方便, 令0≤t 根据古德祥等[19]对地处广东中部的四会市大沙区稻田的调查和稻纵卷叶螟自然种群生命表的研究结果,稻纵卷叶螟的迁入密度为每日百丛1.130头。若稻纵卷叶螟的标准卵量为100粒每雌,去除雄虫因素后第2代、第3代稻纵卷叶螟成虫的平均卵量为71.8粒和54.6粒。稻纵卷叶螟第2,3代卵至成虫期的日均存活率可分两种情况进行设定:一是根据稻纵卷叶螟自然种群生命表的研究结果[19],第2,3代卵至成虫期的日均存活率分别为0.007 8,0.005 4;二是由稻纵卷叶螟实验种群生命表的研究结果[20],当平均温度25 ℃时,第2,3代卵至成虫期的日均存活率分别为0.009 9和0.007 9。假设稻纵卷叶螟第2,3,4代成虫的日均自然死亡率分别为0.006,0.007和0.005。在0≤t 表1 不同情景下稻纵卷叶螟-天敌系统的参数值1) 如图3-4所示,6种情景下稻纵卷叶螟和天敌种群密度的动态曲线各不相同。图3:A1-A2描述的是在稻纵卷叶螟迁出率、天敌种群不变的条件下,天敌对稻纵卷叶螟卵至蛹期的控制作用存在与否对系统动态变化所产生的影响。图3:A3-A4描述的是在稻纵卷叶螟迁出率不变的情况下,天敌的存活率提高10倍,天敌对稻纵卷叶螟卵至蛹期的控制作用存在与否对系统所产生的影响。4种模拟情景中,第2,3,4代稻纵卷叶螟种群密度峰值分别出现在水稻移栽后第18-19天,第44-45天和第69-70天。情景1、情景3中稻纵卷叶螟第2,3代的种群密度均高于情景2、情景4,而第4代种群密度却低于情景2和情景4。情景1中第2代稻纵卷叶螟种群密度最高,为1.237头/每日百丛。4种情景下天敌种群密度随时间的推移均呈现明显的上升趋势,峰值出现在水稻移栽后第84-86天(图3:B1-B4)。可以看出,在第2代稻纵卷叶螟持续迁入期间,由于稻田天敌对稻纵卷叶螟卵至蛹期的控制作用,第4代卷稻纵叶螟成虫的密度将有明显减少,峰值将比第2代降低40.54%(图3:A1)。反之,若不考虑天敌的作用,第4代稻纵卷叶螟成虫密度的峰值仅比第2代降低17.20%(图3:A2)。情景3和情景4中,若采取适当的措施保护天敌(如不施用杀虫农药等),将稻田天敌的存活率提高10倍,则天敌种群密度的增长比情景1和情景2更为明显(图3:B3-B4)。情景3中天敌种群密度最高,达每日百丛4.066头(图3:B3)。此时,如果不考虑卵至蛹期天敌的控制作用,第四代稻纵卷叶螟成虫的密度峰值达每日百丛0.773头,比有天敌控制时的密度值高18.02%。与情景1相比,第四代稻纵卷叶螟成虫的密度峰值也略有上升,说明如果不考虑卵至蛹期天敌的控制作用,即使提高天敌的存活率,仍然无法阻止稻纵卷叶螟在水稻生长中后期发生为害。 图4:A5-A6描述的是在天敌的自然死亡率增加10倍、迁飞率增加近1倍的情况下,天敌对稻纵卷叶螟卵至蛹期的控制作用存在与否对系统动态变化所产生的影响。与前4种模拟情景相比,这两种模拟情景下第2,3,4代稻纵卷叶螟种群密度的峰值均有不同程度的增加,分别出现在水稻移栽后第20-21天,第47-48天和第75-76天,与前4种情景相比略显滞后。情景5中天敌对稻纵卷叶螟存在一定的控制作用,故尽管稻纵卷叶螟第2,3代的种群密度均高于情景6,但第4代种群密度却低于情景6。在天敌种群密度较大幅度减少时,如果不考虑其对稻纵卷叶螟的控制作用,稻纵卷叶螟在水稻生长中后期密度均显著上升,第四代稻纵卷叶螟的种群密度最高可达2.935头/每日百丛,比第2代增加了近2.5倍(图4:A6)。图4:B5-B6中显示,两种模拟情景下,天敌种群的密度随时间小幅波动,峰值出现在水稻移栽后第83-85天。随着稻纵卷叶螟继发代成虫大量迁出稻田,天敌种群密度开始下降。情景6中的天敌种群密度比情景5略低,为每日百丛1.655头,显著低于前4种模拟情景。 表2 STELLA模型中不同世代稻纵卷叶螟成虫动态变化的方程1) 图3 不同情景下稻纵卷叶螟成虫和天敌种群密度的动态变化(Ⅰ) 从图3:A1-A4的4种模拟情景来看,这4种情景下稻纵卷叶螟种群密度峰值的总体趋势表现为从迁入代、盛发代到继发代依次递减。这是由于在第2代稻纵卷叶螟持续迁入田间时,正值水稻分蘖阶段,在当地适宜的温、湿度配合下,稻纵卷叶螟进入了快速生长发育期,而稻田的天敌种群处在开始增长的阶段,其密度低于其它各世代发生期的密度,对稻纵卷叶螟的控制作用也较弱,因而稻纵卷叶螟第2世代的日均存活率相对较高,种群密度在早稻全育期为最高。在第4代发生期,因为气温高、湿度低,对稻纵卷叶螟的生长发育产生了不利影响,同时稻田天敌种群不断增殖,对稻纵卷叶螟的控制作用明显提高。当早稻绝大部分已转入黄熟阶段,第4代稻纵卷叶螟幼虫难以取食为害,死亡率增加,蛹化成虫后,陆续迁飞出稻田。在第3代发生时期,日平均相对湿度与日平均温度与第2代发生时期大体相同,但这一阶段受降雨的影响较大,稻纵卷叶螟的日均存活率下降。此时的天敌种群密度介于第2代、第4代时的种群密度之间,故第3代稻纵卷叶螟的失踪率和种群密度也处于两者之间。 图4:A5-A6的2种模拟情景下,稻纵卷叶螟种群密度峰值的总体趋势均表现为从迁入代、盛发代到继发代依次递增,与前4种情景正好相反。由于田间天敌密度处于较低水平,对害虫的控制作用较弱,使得第2代稻纵卷叶螟大量增殖,第3,4代虫口密度上升,呈暴发为害的态势。说明一定数量天敌的存在,对有效地防治和控制稻纵卷叶螟的发生与发展有着重要意义。根据古德祥[15]对稻纵卷叶螟自然种群生命表的研究结果,捕食性天敌的捕食作用是第2代稻纵卷叶螟种群一龄幼虫失踪死亡的关键因素,3~5龄幼虫的“失踪”是影响稻纵卷叶螟全年种群数量的关键因子,如果不能保护和利用天敌,充分发挥天敌的控虫效能,将会给稻纵卷叶螟种群创造滋养生息的机会,增加了第3,4代稻纵卷叶螟的防治压力。如能在虫害发生初期,适量补充一定数量的天敌以扩大田间种群发展的基数,可提高对水稻中后期稻纵卷叶螟的控制效能。 影响稻纵卷叶螟自然种群死亡的因素很多,各种因素造成的死亡率在不同世代的不同虫态均有差异。本研究未单独考虑气候、田间管理(如施用农药)等因素的影响,但采用的参数如第2、3代卵至成虫期的日均存活率等,实际上是气候因素、天敌捕食和寄主在不同发育阶段综合作用的结果,在分析时可加以综合考虑,对稻纵卷叶螟和天敌系统的动态进行合理的判断和解释。另一方面,研究重点关注的是稻纵卷叶螟不同世代与天敌系统的动态变化规律,故在模拟过程中未对稻纵卷叶螟种群不同虫态之间的数量转化关系及影响因素进行详细分析,尽管用稻纵卷叶螟卵至成虫期的日均存活率来代替不同虫态的存活率,对结果可能会产生一定的影响,但这一处理方法便于以天为时间单位进行模拟,从而克服由于各虫态历期不同,以虫态划分年龄组造成的不等距问题,从系统分析的观点来看,对结果的影响也是极为有限的。 本研究将系统动态模拟软件STELLA引入到害虫种群生态学的研究中,在对稻田害虫种群进行逐日动态模拟的过程中,通过软件的可视化环境对模型结构、运行参数和运行结果进行调整修正,使其更适合于对稻纵卷叶螟—天敌系统相互依存、相互作用的复杂问题开展情景分析和控制策略的研究。在模拟中发现,在第2代稻纵卷叶螟持续迁入期间,稻田天敌对稻纵卷叶螟卵至蛹期的有效控制,将会使第4代卷稻纵叶螟成虫的密度明显减少,如果能在虫害发生初期,适量补充一定数量的天敌以扩大田间种群发展的基数,可提高对水稻中后期稻纵卷叶螟的控制效能。 参考文献: [1]WATT K E F.Mathematical models for use in insect pest control[J].Canadian Entomologist Suppl,1961,19(1):62. [2]WATT K E F.Mathematical population models for five agricultural crop pests[J].Memoirs of the Entomological Society of Canada,1963,32:83-91. [3]MORRIS R F.Predictive population equation based on key factors[J].Memoirs of the Entomological Society of Canada,1963,32: 16-21 [4]HUFFAKER C B.Some aspects of assessing efficiency of natural enemies[J].Canadian Entomologist,1976a,101(3/4):425-447. [5]HUFFAKER C B,MESSENGER P S.Theory and Practice of Biological Control[M].New York: Academic Press,1976. [6]丁岩钦,陈玉平.系统分析及其在生态学中的应用[J].生态学杂志,1982,3 (1) : 46 -51. [7]李典谟,王景明.确定动态经济阈值的微计算机模型[J].生态学报,1986,6 (1) : 53 -59. [8]尹长民,王洪全.系统工程与害虫防治[J].系统工程,1984,2 (1) : 9-14. [9]庞雄飞.昆虫种群系统的研究概述[J].生态学报,1990,10(4):373-378. [10]庞雄飞,梁广文.害虫的种群系统控制[M].广州:广东科技出版社,1995. [11]徐汝梅,成新跃.昆虫种群生态学——基础与前沿[M].北京:科学出版社,2005. [12]王其藩.系统动力学(修订版)[M].上海:上海财经大学出版社,2009. [13]钟永光,贾晓菁,李旭.系统动力学[M].北京:科学出版社,2009. [14]李旭.社会系统动力学——政策研究的原理、方法和应用[M].上海: 复旦大学出版社,2009. [15]贾仁安,丁荣华.系统动力学: 反馈动态性复杂分析[M].北京:高等教育出版社,2002. [16]钟承茂.稻纵卷叶螟的为害症状与发生规律[J].农技服务,2008,25(1):40,60. [17]王 星,曾爱平.基于系统科学的害虫种群系统控制研究[J].广西植保,2009,22(3):24-26. [18]何淼,王寿松.稻纵卷叶螟-捕食性天敌模拟系统研究[J].昆虫天敌,1990,12(4):151-157. [19]古德祥,周昌清,汤鉴球.稻纵卷叶螟自然种群平均生命表[J].中山大学学报:自然科学版,1989,8(8): 37-43. [20]张润杰,古德祥.稻纵卷叶螟实验种群模拟模型[J].中山大学学报:自然科学版,1989,8(8):29-36.1.3 参数赋值

2 结果分析


3 讨 论
3.1 前4种情景下稻纵卷叶螟种群密度的动态分析
3.2 后两种情景下稻纵卷叶螟种群密度的动态分析
3.3 稻纵卷叶螟种群密度动态的影响因素
4 结 论
