米氏冰草营养繁殖潜在种群与现实种群的关系
2012-04-12金晓明刘及东卢欣石孙兆军
金晓明,刘及东,卢欣石,孙兆军
(1.呼伦贝尔学院生命科学学院,内蒙古 呼伦贝尔021008;2.北京林业大学草地资源与生态实验室,北京100083;3.宁夏大学新技术应用研究开发中心,宁夏 银川750021)
*芽库是营养繁殖的潜在种群,种子库是有性繁殖的潜在种群,二者共同组成了植物的繁殖库[1]。对于草本植物,芽既是重要的越冬构件,又是扩大种群的重要指标。芽进一步发育,可能形成新的苗和分株,也可能形成新的根茎,是无性系空间生态位的扩展和持续更新的基础[2,3]。通常芽库和苗库被称为无性系植物无性繁殖的潜在种群,分株被称为无性繁殖的现实种群。
当地上植被遭受灾难性破坏后,芽库能够使种群迅速重建,大大减少了物种灭绝的几率[4,5]。在植物群落中,许多草本植物群落的成功建植很少依赖种子,而是依赖营养繁殖方式。多年生草本植物具有较低的有性繁殖率[6],特别在天然草地上,根茎型禾本科植物靠种子成功定居的比率很小,据统计,其数量比例占不到总地上分株数量的6%[7,8]。多年生草本植物能使其地上种群通过根茎上的分蘖节或芽库形成新的地上枝条,对种群结构和季节动态起着关键作用[9]。植物的营养繁殖对种群的空间结构及其动态等具有较大影响,因此了解营养繁殖潜在种群对现实种群的贡献是必要的。在一些生态系统中,芽库在植物局域种群的持续更新和维持生存、群落对不利环境、种间竞争、取食压力、植被结构和生产力等方面具有重大意义,甚至起着决定性作用[10]。例如,在北美高草草原,芽库是调控地上植被净初级生产力的最主要来源[11],原因是99%以上的地上茎秆由芽库形成[12,13]。
植物通过调控芽库中芽的输入、输出及密度等,对环境干扰作出反应,进而调控着植物种群的组成和动态[14]。在一些极端生境,植物会放弃有性繁殖方式[15],完全以芽库作为潜在种群完成种群的更新。因此,芽库的动态变化对种群、群落及生态系统的研究具有重要意义。目前有关芽库分类的研究相对较少。杨允菲[16]根据芽的着生位置分为根茎芽和分蘖节芽,根茎芽是指由根茎节间和根茎顶端长出的芽,分蘖节芽为母株分蘖节上长出的芽。Kline和Klineová[17]根据芽着生器官的不同将芽分成21类,如块根、块茎、鳞芽等。有的根据芽的发生时间分为1年生芽(annual buds)和多年生芽(perennial buds)[18]。Klimeová和 Klimě[19]根据芽数、季节动态、扩散能力、休眠方式等将芽库分为3类:永久芽库、季节性芽库和潜在芽库。
国内外有关种子库的进化生态学的研究很多[20,21],关于芽库的研究主要集中在其重要作用方面,而关于芽库的繁殖方式、动态及结果方面的研究却很少[22-24]。植物个体的生长被认为是构件数量动态变化过程,枝系伸展实际上是芽库出生率和死亡率的统计学过程[9]。在调查了银桦(Betulapendula)芽的命运的基础上,并用矩阵模型分析了芽库的增长及数量动态[25,26]。另外对芽库的研究方法上,还引用了三角函数[27],马尔可夫(Markov)模型[28],和矩阵的灵敏度分析等[29]。但国内的研究则刚刚起步。
本研究以沙地不同生境的米氏冰草(Agropyronmichnoi)为对象,分析其生活史过程中芽库与分株的关系,揭示米氏冰草营养繁殖潜在种群与现实种群的关系,为进一步研究无性系植物的繁殖规律及扩展机理提供理论依据。
1 材料与方法
1.1 研究地自然概况
研究区位于中国呼伦贝尔草原境内。地理位置为东经11°44′~118°54′,北纬49°08′~49°11′,海拔588.4~618.0 m。具温带大陆干旱半干旱过渡气候特征。年均气温-1.5℃,降水量230~300 mm(70%集中于夏秋季),而蒸发量为降水量的4~6倍;年均风速4.5 m/s,最大风速为20 m/s,全年8级以上大风日数平均30 d以上。地带性植被为以贝加尔针茅(Stipabaicalensis)和羊草为优势种的群落。沙漠化草地的植被主要以草本植物种为主,其中有米氏冰草、东北牡蒿 (Artemisiajaponicavar.manshurica)、扁蓄豆(Melissitusruthenius)、狗尾草(Setariaviridis)、百里香(Thymusvulgaris)、猪毛菜(Salsolafoliosa)、沙蓬(Agriophyllumsquarrosum)、沙米(Agriophyllumarenarium)等。另外,还零星分布着少量的灌木,如差不嘎蒿(Artemisiahalodendron)和小叶锦鸡儿(Caraganamicrophlla)等。
1.2 研究方法
本研究是在流动沙地(P1)和固定沙地(P2)内进行。采用营养繁殖潜在种群和现实种群的调查方法:在2009年10月2日、2010年6月14日、7月7日、8月1日和9月17日,采用单位面积挖掘取样法,面积大小为25 cm×25 cm,深度为30 cm,10个重复,将四周的土挖开使样方成土柱状,连同地上部分一并装入样袋中,尽量保持土柱的完整性。回室内用水轻轻冲洗,保持所有地上和地下部分的自然联系,以便于鉴别。
按每个样方计数以下内容:1)同一龄级的分株、苗、芽和根茎等构件。2)根茎芽和根茎苗、分蘖节芽和分蘖节苗。3)春性分株和冬性分株(6月)。
1.3 样品的处理与分析
冬、春性分株的鉴定方法:在春季各分株最下部的叶片尚未开始黄化以前进行。具体鉴别方法是:凡是分蘖株下部留有枯叶鞘、枯叶片、枯叶尖,或留有任一干枯残痕的分蘖株均为冬性分株,其他于本生长季出生的分株为春性分株[30]。
冬性芽和冬性苗的鉴定方法:一般无性系在生长季末期形成的芽和苗分别以冬性芽和冬性苗的形式越冬,并在次年继续生长,冬性芽发育为春性分株,冬性苗发育为冬性分株[2]。
1.4 数据处理
营养繁殖潜在种群转化为现实种群的效率=下一年现实种群的总数量/当年生长季末期形成的潜在种群数量。
数据采用Excel软件和SPSS l6.0软件处理。将调查数量指标均换算成1 m2的数据指标。数据统计,采用ANOVA的Duncan法进行方差分析,取各样方的平均值(M)及标准差(SD)。
2 结果与分析
2.1 营养繁殖潜在种群与现实种群的关系
固定沙地的营养繁殖潜在种群的总数量显著高于流动沙地,是流动沙地的2.9倍(表1)。米氏冰草营养繁殖潜在种群由冬性苗和冬性芽组成,且冬性苗的比率比冬性芽高3~4倍。表明,在生长季末期,米氏冰草已形成大量的营养繁殖潜在种群,且大部分以冬性苗的形式越冬。另外,流动沙地的冬性苗比率大于固定沙地,固定沙地的冬性芽比率大于流动沙地。表明,环境干旱可使米氏冰草的生育期缩短。2010年6月中旬,为米氏冰草的抽穗期,固定沙地的总分株数量显著高于流动沙地,是流动沙地的4.4倍;固定沙地的营养分株的数量显著高于流动沙地,是流动沙地的6.7倍。米氏冰草的分株由冬性分株和春性分株组成,其中冬性分株的比率达98.9%以上,春性分株的比率低于2.0%。冬性分株中包括生殖分株和营养分株,且流动沙地和固定沙地营养分株的比率分别为58.5%和89.1%,生殖分株的比率分别为40.4%和10.6%。春性分株中全部为营养分株。表明,生长季初形成的米氏冰草营养繁殖现实种群主要是冬性营养分株,不同生境营养分株的比率总是高于生殖分株的比率;环境干旱可促使生殖分株的比率增高。
营养繁殖潜在种群与现实种群之间的数量转化效率见表2,总营养繁殖潜在种群转化为总现实种群的效率,在流动沙地和固定沙地分别为38.3%和58.2%,其中冬性苗转化为冬性分株的效率分别为48.0%和84.4%,冬性芽转化为春性分株的效率分别为2.0%和0.4%。表明,在沙地不同生境,米氏冰草的营养繁殖潜在种群与现实种群的转换中,冬性苗的贡献远远大于冬性芽。另外,冬性苗转化为冬性生殖分株的效率分别为19.4%和9.0%,冬性苗转化为冬性营养分株的效率分别为28.1%和75.2%,冬性芽转化为春性营养分株的效率分别为2.0%和0.5%。表明,在沙地生境,营养繁殖潜在种群转化为营养分株的效率大于转化为生殖分株的效率,即营养繁殖为米氏冰草的主要繁殖方式。
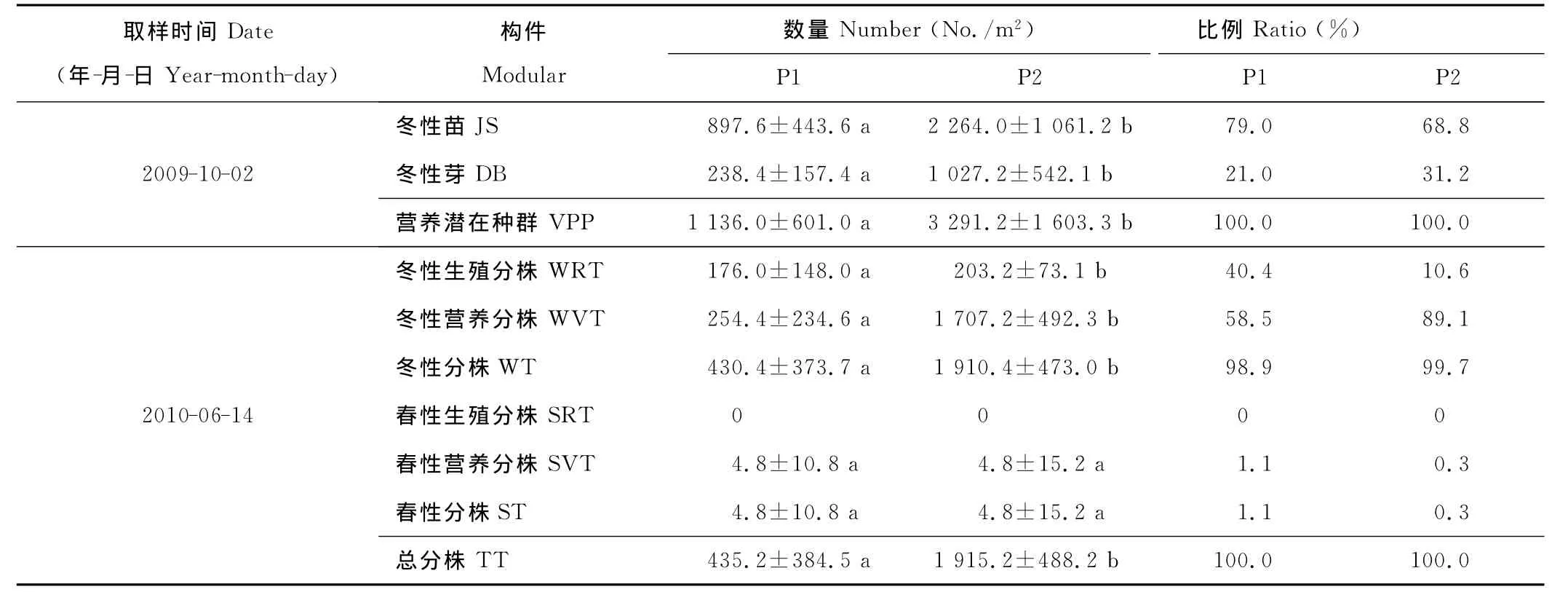
表1 米氏冰草营养繁殖潜在种群与现实种群的数量特征Table 1 The quantitative character of potential population and reality population of vegetative reproduction of A.michnoi
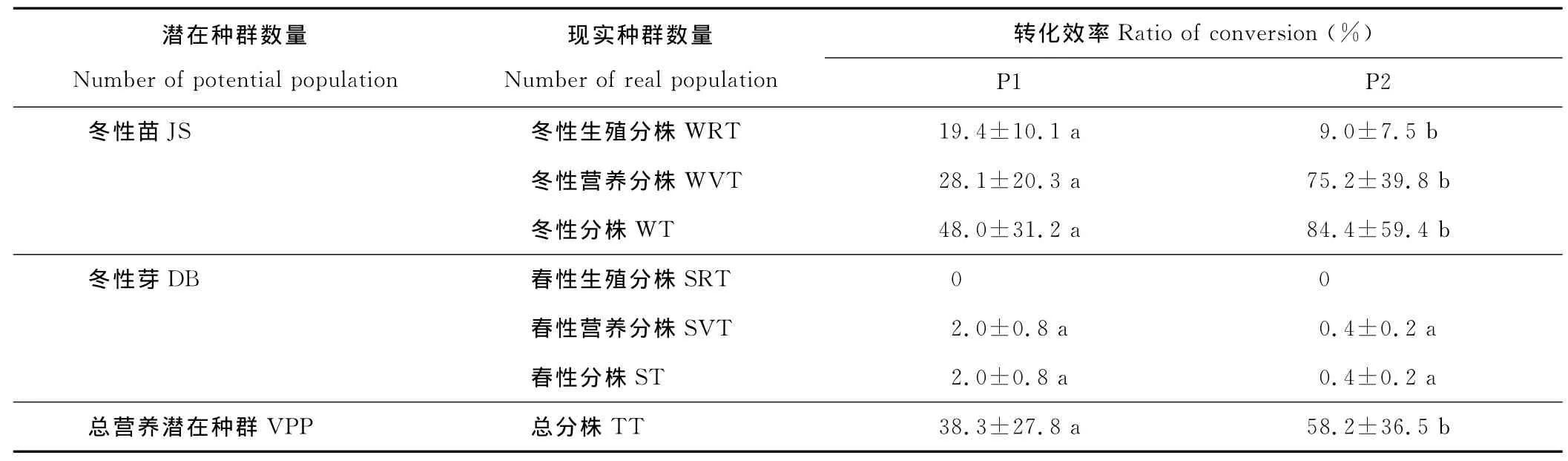
表2 营养繁殖潜在种群与现实种群之间的数量转化效率Table 2 Efficiency of changes on potential population and reality population of vegetative reproduction of A.michnoi
2.2 营养繁殖潜在种群与现实种群年龄结构之间的转化效率
在2009年10月,即米氏冰草的生长季末期,冬性苗包括根茎苗和分蘖节苗(表3)。其中,固定沙地的根茎苗和分蘖节苗分别比流动沙地的高1.5和4.1倍。在冬性苗的比例上,流动沙地的根茎苗∶分蘖节苗=60.5∶39.5,固定沙地的根茎苗∶分蘖节苗=36.3∶63.7。表明,流动沙地的根茎苗比率高于固定沙地,固定沙地的分蘖节苗比率高于流动沙地。2010年6月,即次年米氏冰草各龄级分株数量组成上,在流动沙地以1龄级占优势,并随着龄级的增加呈减少趋势。在固定沙地以2龄级占优势,其次是1,3和4龄级。固定沙地的1,2,3和4龄级分株数量分别比流动沙地高2.7,6.0,9.8和7.0倍。表明,米氏冰草的各龄级分株数量,固定沙地均高于流动沙地,且龄级越高,差距越大。
冬性苗与次年现实种群各龄级分株之间的数量转化效率见表4,可以看出,冬性苗转化为营养繁殖现实种群的效率,在流动沙地和固定沙地分别为48.4%和84.7%;根茎苗转化为1龄级分株的效率,在流动沙地和固定沙地分别为45.4%和81.5%;分蘖节苗转化为2~4龄级分株的效率,在流动沙地和固定沙地分别为52.9%和86.6%。流动沙地和固定沙地的米氏冰草的冬性苗中,分蘖节苗转化为现实种群的效率比根茎苗分别高1.2和1.1倍。根茎苗与分蘖节苗的转化效率,固定沙地分别是流动沙地的1.8和1.6倍。表明,米氏冰草的营养繁殖现实种群主要由前一年的冬性苗发育形成。由分蘖节苗形成分株是一种消耗能量相对较小的营养繁殖方式,而由根茎苗形成分株是一种消耗能量相对较大的营养繁殖方式。适宜生境中冬性苗的转化效率高于不利生境。
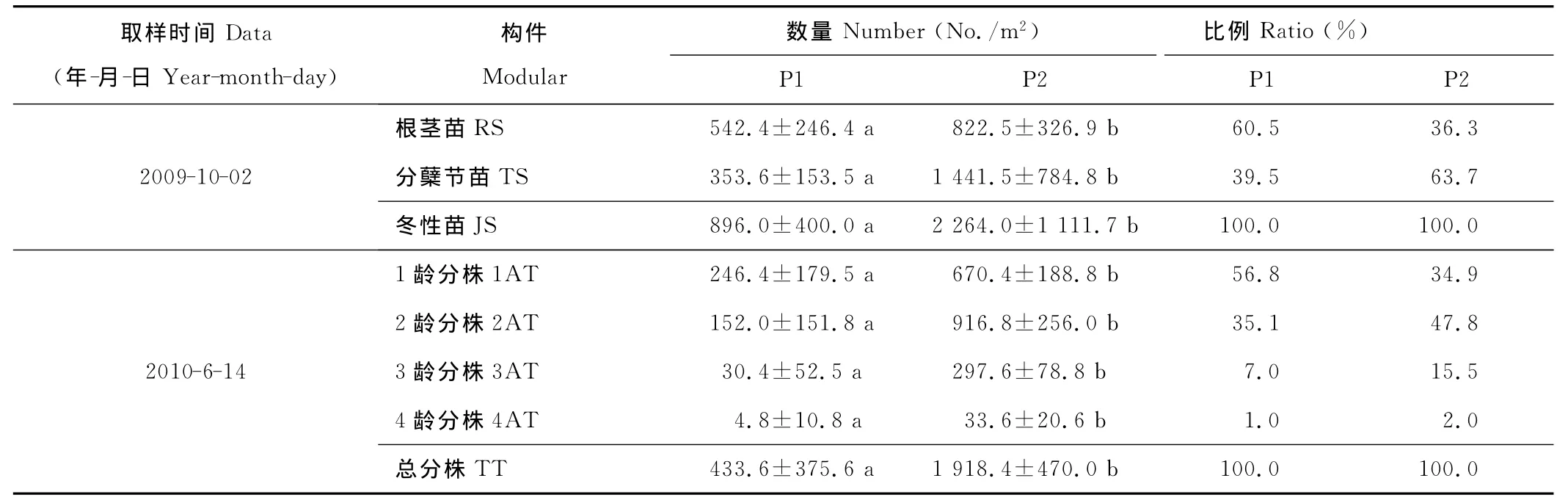
表3 米氏冰草营养繁殖潜在种群与现实种群中各龄级分株的数量特征Table 3 The quantitative character of potential population and tillers of every age class in reality population of A.michnoi

表4 冬性苗与现实种群中各龄级分株之间的转化效率Table 4 The Efficiency of changes on dominant seedlings and tillers of every age class in reality population of A.michnoi
3 讨论与结论
在草地生态系统中,多年生草本植物个体的繁殖潜力可通过构件在数量和大小上如何生长来体现[31-34],其中芽库数量的多少对净初级生产力具有直接影响[11]。本研究表明,随着沙地植被的恢复,米氏冰草营养繁殖潜在种群(苗和芽)的总数量呈增长趋势,即固定沙地的营养繁殖潜在种群比流动沙地高2.9倍。另外,冬性苗比冬性芽高3~4倍。表明,在生长季结束前,大部分营养繁殖的潜在种群已长成苗,保证潜在种群较高的越冬率,并为下一年种群的存活及增长做了准备。这也是米氏冰草营养繁殖潜在种群对环境适应的一种表现。
在生长季前期的6月,营养繁殖现实种群中99% 以上是由前一年的冬性苗转化形成的冬性分株,只有1%左右是由前一年的冬性芽转化形成的春性分株。这一结果与松嫩平原野大麦冬眠构件的研究基本相似,即在生长季前期的6月初,野大麦种群的分株中有95%是由冬眠苗形成的冬性分株,只有5%是由冬眠芽形成的春性分株[30]。表明,米氏冰草的现实种群基本来源于冬性苗。大部分米氏冰草的冬性芽没有在第2年形成新的分株,这有可能是因为米氏冰草的冬性芽在冬季寒冷气候条件下,因养分积累不足而冻死,关于这一点有待于进一步的研究。
在冬性苗中包含根茎苗和分蘖节苗,根茎苗转化为1龄级分株的效率,在流动沙地和固定沙地分别为45.4%和81.5%;分蘖节苗转化为2~4龄级分株的效率,在流动沙地和固定沙地分别为52.9%和86.6%。表明,随着沙地植被的恢复,米氏冰草营养繁殖潜在种群转化为现实种群的效率明显提高。在根茎苗和分蘖节苗中,由分蘖节苗形成分株比由根茎苗形成分株相对更容易,即能耗较低。前者可能的原因是,流动沙地的根茎苗或分蘖节苗露出地面,进行其营养生长过程中,要抵抗恶劣环境,因此消耗比固定沙地更多的能量才能生存下来。另外,在大量营养繁殖潜在种群中,到第2年还能存活的比例不是很大,这可能与米氏冰草在冬眠之前根茎的养分贮存量不足,以及在次年幼苗生长过程中要消耗大量根茎贮存的养分,所以在众多幼苗的养分竞争中,使有一部分幼苗因得不到最基本的养分而导致死亡。因此,流动沙地的米氏冰草的营养繁殖策略与R-对策者的理论是相吻合的[35],而固定沙地的米氏冰草的营养繁殖策略与K-对策者的理论是相吻合的[35]。后者可能的原因是,根茎苗直接生长在根茎顶端或节间,在发育为新分株时,虽然可以从根茎直接获取充足的养分,但要占据一个新的生态位空间,就必须消耗大量的能量才能从土壤中破土而出。而分蘖节苗着生在分株上,可以直接获取分株的养分,但形成新分株时可顺着其着生的分株的茎而生长,因此不需要消耗太多能量。
[1] Abernethy V J,Willby N J.Changes a long a disturbance gradient in the density and composition of propagule banks in floodpla in aquatic habitats[J].Plant Ecology,1999,140(2):177-190.
[2] 杨允菲.松嫩平原赖草无性系生长及其构件的年龄结构[J].应用生态学报,2004,15(11):2109-2112.
[3] 李海燕,李建东,徐振国,等.内蒙古图牧吉自然保护区羊草种群营养繁殖特性的比较[J].草业学报,2011,20(5):19-25.
[4] Lee P.The impact of burn intensity from wildfires on seed and vegetative banks,and emergent understory in aspendom inated boreal forests[J].Canadian Journal of Botany,2004,82:1468-1480.
[5] Liu W,Zhang Q,Liu G.Seed banks of a river-reservoir wetland system and their implications for vegetation development[J].Aquatic Botany,2009,90(4):7-12.
[6] 王桂芹,高瑞如,王玉良,等.异质生境空心莲子草的结构基础与生态适应性[J].草业学报,2011,20(4):143-152.
[7] Benson E J.Effects of fire on tallgrass prairie plant population dynamics[A].Thesis[M].Manhattan USA:Kansas State U-niversity,2001:23-29.
[8] 金晓明.米氏冰草繁殖生态学特征及种群竞争机制[D].北京:北京林业大学,2011.
[9] Harper J L.Population Biology of Plants[M].London:Academic Press,1977:15-23.
[10] Hartnett D C,Setshogo M P,Dalgleish H J.Bud banks of perennial savanna grasses in Botswana[J].African Journal of Ecology,2006,(44):256-263.
[11] Knapp A K,Smith M D.Variation among biomes in temporal dynamics of aboveground primary production[J].Science,2001,291:481-484.
[12] Benson E J,Hartnet D C,Mann K H.Below ground bud banks and meristem limitation in tallgrass prairie plant populations[J].American Journal of Botany,2004,91:416-421.
[13] Benson E,Hartnett D.The role of seed and vegetative reproduction in plant recruitment and demography in tallgrass prairie[J].Plant Ecology,2006,187:163-178.
[14] Dalgleish H J,Hartnett D C.The effects of fire frequency and grazing on tallgrass prairie productivity and plant composition are mediated through bud bank demography[J].Plant Ecology,2009,201:411-420.
[15] Eckert C G,Dorken M E,Mitchell SA.Loss of sex in clonal populations of a flowering plant,Decodon verticillatus(Lythraceae)[J].Evolution,1999,53:1079-1092.
[16] 杨允菲.松嫩平原几种根茎型禾草种群的营养繁殖特性及其持续更新分析[J].草业学报,1996,5(2):43-48.
[18] Tolvanen A,Schroderus J,Henry G H R.Age-and stage-based bud demography ofSalixarcticaunder contrasting muskox grazing pressure in the High Arctic[J].Evolutionary Ecology,2001,15(6):443-462.
[20] Fenner M.Seed Ecology[M].London:Chapman and Hall,1985:66-113.
[21] Baskin C C,Baskin J M.Seeds:Ecology,Biogeography,and Evolution of Dormancy and Germination[M].San Diego,California,USA:Academic Press,1998.
[22] Busso C A,Mueller R J,Richards J H.Effects of drought and defoliation on bud viability in two caespitose grasses[J].Annals of Botany,1989,63:477-485.
[23] Tuomi J,Nilsson P,Astrom M.Plant compensatory responses:bud dormancy as an adaptation to herbivory[J].Ecology,1994,75:1429-1436.
[24] Hendrickson J R,Briske D D.Axillary bud banks of two semiarid perennial grasses:occurrence,longevity,and contribution to population persistence[J].Oecologia,1997,110:584-591.
[25] Maillette L.Structural dynamics of silver birch 1.the fates of buds[J].Journal of Applied Ecology,1982,19(7):203-211.
[26] Maillette L.Structural dynamics of silver birch 2.a matrix model of the bud population[J].Journal of Applied Ecology,1982,19(8):219-231.
[27] Maillette L.Seasonalmodel of modular growth in plants[J].Journal of Ecology,1992,80(2):123-130.
[28] Maillette L.The value of meristem states,as estimated by a discrete time Markov chain[J].Oikos,1990,59(5):235-241.
[29] Lehtila K,Tuomi J,Sulkinoja M.Bud demography of mountain birchBetulapubescensspp.Tortuosa near tree line[J].Ecology,1994,75(4):945-951.
[30] 杨允菲,郑慧莹,李建东.根茎禾草无性系种群年龄结构的研究方法[J].东北师范大学学报,1998,1(1):49-53.
[31] Harper J L.The concept of population in modular organisms[A].In:May R M.Theoretical Ecology,Principles and Applications.(2nd ed)[M].Oxford:Blackwell Scientific Publications,1981:53-77.
[32] Watson M A,Casper B B.Morphogenetic constraints on patterns of carbon distribution in plants[J].Annual Review of Ecology and Systematics,1984,15(7):233-258.
[33] Fox J F.Shoot demographic responses to manipulation of reproductive effort by bud removal in a willow[J].Oikos,1995,72:283-287.
[34] Preston K A.Architectural constraint on flower number in a photoperiodic annual[J].Oikos,1998,81:279-288.
[35] Mac Arthur R H,Levins R.The limiting similarity,convergence and divergence of coexisting species[J].American Naturalist,1967,101(5):337-385.
