大气CO2富集对根际NaCl盐渍的人参果植株干物质生产和水分利用的影响
2012-01-30陈凯胡国谦KEUTGENNorbertKEUTGENAnnaJANSSENSMarcLENZFritz
陈凯,胡国谦,KEUTGEN Norbert,KEUTGEN Anna,JANSSENS Marc,LENZ Fritz
(德国波恩大学果树与蔬菜研究所,德国波恩市D-53121)
0 引言
作物生长和发育的立地环境(如大气、水分和土壤等条件)深刻影响植株干物质生产和水分利用效率,进而制约作物的经济产量和产品质量。环境因素不仅直接或间接调控植株干物质生产、积累和分配,而且左右植物吸收和利用水分的效率(Bazzaz,1990;Idso and Idso,1994;Piao et al.,2010)。农田土壤和灌溉水分中累积NaCl是当前世界上最常见的盐渍化问题。农用灌溉水中NaCl质量浓度大于20 mg·L-1会显著胁迫作物根系生理活动功能,影响土壤水分和养分有效性,限制作物生长发育和降低生产力(Matar et al.,1975;Zeroni and Gale,1989;Grattan,2002;García-Sánchez and Syvertsen,2006)。大气CO2富集一方面是全球气候变化的主要动因,另一方面能促进作物生长发育和提高生产力,这是因为CO2是植物光合作用的必需原料,而光合作用是作物干物质生产的基础(Eamus and Jarvis,1989;Bazzaz,1990)。在农业生产中,作物根际土壤和水分盐渍及大气CO2富集,对植株生长发育和生理代谢的复合互作影响比它们各自的单独作用要复杂得多。而且,这种现象普遍存在。因此,有必要研究作物在根际NaCl盐渍和大气CO2富集情况下的生理生化反应及其生物学表现与适应机理,深入探讨植物形态结构和生态生理变化及NaCl盐渍和CO2富集对植株干物质生产和水分利用的双重影响和互作效应。
人参果(Solanum muricatum Ait.)是原产南美洲的一种短生长期大田园艺作物。其果实美味甘甜、营养丰富,是哥伦比亚、厄瓜多尔、玻利维亚、秘鲁和智利等国鲜果市场上常见果品,也是新西兰、澳大利亚、以色列、北美和欧洲诸国引入栽培或进口的水果(Chen et al.,1999a)。研究发现,该作物根际土壤盐渍和大气CO2富集不仅影响植株营养生长和生殖生长(Chen et al.,1999a),而且制约叶片光合作用功能和气体交换(Chen et al.,1999b)。
本研究基于下列科学假设:植物被根际盐渍和大气CO2富集诱导的生理反应可能会进一步引起干物质生产效率及其向不同器官中分配的变化(Flore and Lakso,1989;Lenssen et al.,1993)。根际NaCl质量浓度和大气CO2体积分数同时升高,会导致植物叶片气孔关闭和蒸腾作用下降(Chen et al.,1999b),植株水分利用特性的改变是植物对根际NaCl盐渍和大气CO2富集的另一个重要的生理反应(Rozema et al.,1991)。同时,植物叶片水势和光合作用水分利用率也随大气CO2体积分数上升而提高,但又因根际NaCl盐渍加重而下降(Chen et al.,1999b)。据此推断,根际NaCl盐渍和大气CO2富集在大气—植物—土壤—水分的生态系统中有复合互作关系。这种关系又进一步影响植物的干物质生产和水分利用。本研究旨在探讨大气CO2富集对根际受NaCl盐渍胁迫的人参果植株干物质生产和分配及水分利用的影响,同时深入研究它们对该植物上述生理代谢的复合互作效应,为该作物生产提供科学的理论依据和积累有用的实验资料。
1 材料和方法
本研究的模拟试验在德国波恩大学果树与蔬菜研究所进行。取已生根的一月龄人参果栽培品种“Xotus”半木质化扦插苗,栽培于容积为2 L的塑料盆中。每盆栽一株。盆中填满直径为0.7~1.2 mm的石英砂,表面铺一层直径为1~2 cm的石子,以减少培养基质蒸发水分。在整个试验期间,每天下午浇足自来水一次(根据植株大小,每株浇150~200 mL水),并每周浇两次200 mL的Hoagland营养液(Hoagland and Arnon,1938)。营养液的pH值用98%的H2SO4调整至6.0±0.2。供试植株经在田间环境条件下预培养一周后分成两组。一组继续供给未添加NaCl的Hoagland营养液,另一组每周供给添加了25 mg·L-1NaCl的Hoagland营养液两次。考虑到根际NaCl盐渍比大气CO2富集处理对植株干物质生产与分配的影响迟缓,先将经上述处理的供试植株置于田间环境下预处理1个月,再将它们移入可控制环境条件的植物生长培养箱,同时进行上述根际NaCl盐渍和空气CO2富集处理1个月。3个培养箱中空气CO2体积分数控制为(350±10)×10-6、(700±10)×10-6和(1 050±10)×10-6,分别代表自然大气中的CO2水平及其2倍和3倍的水平。供试植株继续定时定量地供给水分和添加或未添加25 mg·L-1NaCl的Hoagland营养液。培养箱中微环境条件为:日/夜光周期16 h/8 h,气温(25±2)℃/(20±2)℃,相对空气湿度70%±5%/80%±5%,空气流通速度(600±20)m3·h-1。白天光周期时用到达植株叶片表层的光合作用光子通量(photosynthetic photon flux)为(550±20)μmol·m-2·s-1的Philips SON-T Agro 400高亚钠光灯照射。培养箱中的CO2体积分数用德产Uras 2-T红外线分析仪(IRGA,Hartmann und Braun AG Meβ-und Regeltechnik,Frankfurt/Main,Germany)监测。在日光周期中,当培养箱内CO2体积分数低于或高于设定水平10×10-6时,即自动补充工业用CO2气体(Technische Gas,Linde AG,Düsseldorf,Germany)或停止供气。在夜间,因植物暗呼吸作用而释放出的剩余CO2由装有10%NaOH溶液的装置通过空气循环流通系统抽除。供试植株在培养箱中放置的位置每周变换一次,以消除不同位置的系统误差及微环境条件的影响。
在每项处理开始及结束时,每项处理各取样5株。叶面积用美产LI-COR 3100面积测量仪(LICOR Inc.,Lincoln,Nebraska,USA)测定,精确度为±0.01 cm2,据此进一步计算单株总叶面积,以及在处理前和后的植株叶面积增长量。所取叶片、枝条、果实、主干和根系样本置于德产TPYL-10冷冻干燥系统中(WKF GmbH,Heistenbach,Germany)干冻至衡重,再称取干重,精确度为±0.01 g。以此计算干物质总增长量和日平均增长速率。植株根/冠比根据每株供试材料地下部分器官与地上部分器官的干重之比求得。将每株试材的干物质增长量除以其叶面积增长量即得其干物质生产效率。干物质向植株各器官中的分配量根据该植株供试前、后各器官中平均增长量来确定。
每天在浇水或供应营养液前和后,分别称取每株试材连同培养基质和盆钵的重量,再扣除在同样条件下无植株的对照盆钵日失水量,以此计算其日耗水量。每株试材的总耗水量即是其在试验期间的日耗水总和。叶片蒸腾系数为植株单位叶面积的平均水分蒸腾量。植株含水量(%)根据其鲜重(fresh weight,WF)和干重(dry weight,WD),用公式(WFWD)/WF×100%求得。根系吸水效率以试验期间植物根系的总干重增长量除以同期植株的总耗水量表示。植株水分利用效率以试验期间干重总增长量除以耗水总量表示。
本研究以根际NaCl盐渍和空气CO2富集两种处理因素进行裂区排列试验设计,分别以根际0 mg·L-1NaCl和350×10-6大气CO2体积分数为两因素的对照。植物生长箱设置350×10-6、700×10-6和1 050×10-6三个水平的CO2体积分数,每个生长箱中放置根际经0 mg·L-1和25 mg·L-1NaCl两种水平的盐渍处理盆栽试材,各处理重复5次。根际NaCl盐渍和空气CO2富集复合处理结果的试验数据,用SPSS统计分析软件(SPSS Inc.,Chicago,USA)进行Tukey-HSD多元变量分析测验。处理效应的显著性以95%的置信度为标准。
2 结果
2.1 叶片生长发育
根际NaCl盐渍和大气CO2富集都影响人参果植株叶片的生长发育,但两者的作用效应存在差异。在本试验期间,单株总叶面积和叶面积增长量因根际受NaCl胁迫而显著减少,但又随大气CO2体积分数的升高而增加(表1)。这些现象表明土壤中NaCl盐渍对植物叶片生长发育有抑制作用,而大气中CO2富集则有促进作用。并且,盐渍对叶片生长发育的抑制作用在CO2体积分数较低时比较高时要显著得多。以上2项指标在25 mg·L-1NaCl处理的植株中分别下降了51%和56%;但与自然空气的处理相比,当CO2体积分数升高1倍及2倍时,它们依次提高了47%、81%及70%和122%(表1)。这些事实说明,根际NaCl盐渍对叶片生长的负作用可由大气CO2富集促进叶片的生长发育而部分地抵消。因此,本试验结束时受根际25 mg·L-1的NaCl盐渍并经大气CO2富集至700×10-6和1 050×10-6双重处理的植株总叶面积和叶面积增长量仅分别增加40%~73%和78%~141%。
植物比叶干重代表其叶片发育质量。根际NaCl胁迫和大气CO2富集均对该指标有改善作用(表1)。在350×10-6CO2下,经25 mg·L-1NaCl盐渍处理和对照植株的比叶干重虽未出现显著差异,但在700×10-6和1 050×10-6CO2的条件下,这项指标相应提高22%和27%。由于根际NaCl盐渍和大气CO2富集的互作关系,在25 mg·L-1NaCl胁迫下,经700×10-6和1 050×10-6CO2处理的植株比叶干重相应增加27%和34%。
2.2 干物质增长
植物器官中干物质增长代表其光合产物的净积累水平。本研究中,供试植株总干物质增加量受根际NaCl胁迫而减少,同时又因大气中CO2升高而增加(表2)。本试验中,植株干物质总量以1 050×10-6CO2且无NaCl盐渍的处理最大、大气350×10-6CO2和根际25 mg·L-1NaCl盐渍的处理最小。在第一个月的盐渍预处理期间,经25 mg·L-1NaCl处理的植株干物质增长量比对照减少25%。经第二个月的根际NaCl盐渍和大气CO2富集复合处理,继续受到NaCl胁迫的植株干物质增长量减少56%;而未经NaCl盐渍且在700×10-6和1 050×10-6CO2条件下生长的植株干物质增长量比对照分别提高79%和133%。由于NaCl胁迫和CO2富集处理的互作效应,前者对植物干物质生产的抑制作用被后者的促进作用所抵消(表2)。因此,在NaCl盐渍的情况下,生长于700×10-6和1 050×10-6CO2条件下的植株干物质总增长量比对照分别提高106%和189%。上述试验结果清楚地表明,大气CO2富集能抵消土壤NaCl盐渍对植物干物质生产的负面作用,从而提高植株的干物质生产力。
植物根/冠比代表植株地下部分器官与地上部分器官生长和发育的协调比例关系。供试植株的该项指标,在根际NaCl盐渍的处理中得以提高,在大气CO2富集的处理中则下降(表2)。这些事实说明,根际NaCl胁迫显著限制植物地上部器官的生长,而同时促进根系的发育。但大气CO2富集不仅能显著促进植物地上部器官生长,而且会有利于根系发育。本试验的第二个月处理期间,经25 mg·L-1NaCl和350×10-6CO2处理的植株根/冠比提高57%;而在无NaCl胁迫条件下,经700×10-6和1 050×10-6CO2处理的植株根/冠比分别下降27%和40%。本试验中,受25 mg·L-1根际NaCl盐渍和700×10-6~1 050×10-6大气CO2富集互作影响的植株根/冠比减小3%~14%。这些事实说明,当根际受NaCl胁迫时,升高大气中CO2体积分数有利于平衡植物地下部和地上部器官生长与发育。
植物干物质生产效率随根际NaCl盐渍化程度和大气CO2升高而显著提高(表2)。这种效应以升高大气CO2水平比加重NaCl胁迫的处理更显著。本试验中在自然空气的CO2条件下,经25 mg·L-1NaCl处理的植株干物质生产效率比未受NaCl胁迫者有提高。未经NaCl盐渍处理的植株,在700×10-6和1 050×10-6CO2下分别比对照提高3%和9%。由于根际NaCl胁迫和大气CO2富集的互作效应,经700×10-6和1 050×10-6CO2处理的植株干物质生产效率相应提高18%和23%。

表1 根际NaCl盐渍和大气CO2富集对人参果植株叶片生长发育特性的影响Table 1Leaf characteristics of pepino plants as affected by NaCl salinity and CO2enrichment
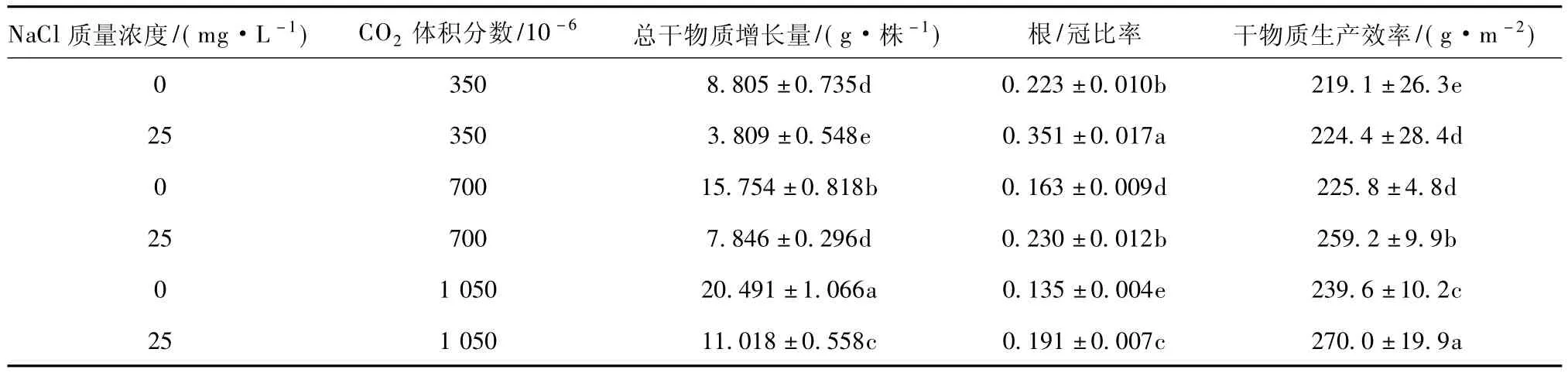
表2 根际NaCl盐渍和大气CO2富集对人参果植株干物质生产的影响Table 2 Dry mass production of pepino plants as affected by NaCl salinity and CO2enrichment
2.3 干物质日增长量
供试植株及其不同器官的干物质日增长量因根际NaCl胁迫而显著下降,同时又由于大气CO2富集而明显上升(表3)。本试验所有处理中,供试植株不同器官的干物质日增长量大小顺序依次为叶片>果实>枝条>根系>主干。这种趋势又以叶片和果实最显著,证明在试验期间这两种器官分别是植株最旺盛的营养生长和生殖生长主导中心,同时说明植株中处于生理代谢最活跃的器官对外界环境的变化(无论是根际NaCl盐渍,还是大气CO2富集)反应最敏感。叶片是植物进行光合作用,制造有机干物质的主要场所(有机碳源);果实是植株利用光合产物,形成经济产品的重要基地(有机碳库)。它们都是调控植株生理代谢的核心器官。因此,它们与植株干物质增长有关的生理生化代谢过程,最容易随环境因素的变化而改变。
在本研究中还观察到,当大气CO2体积分数从350×10-6上升到1 050×10-6时,植株各器官中干物质日增长量呈稳步增长趋势。这种情况在叶片、果实和枝条中表现很明显(表3)。在根际无NaCl盐渍的条件下,经700×10-6和1 050×10-6CO2处理的植株,比对照的总干物质日增长量平均提高79%和133%;而根际在25 mg·L-1NaCl盐渍的情况下,大气CO2为350×10-6、700×10-6和1 050×10-6时,植株干物质增长量依次下降57%、50%和46%。由于根际NaCl盐渍和大气CO2富集的互作效应,经25 mg·L-1NaCl和700×10-6及1 050×10-6CO2处理的植株总干物质增长量则比无NaCl胁迫和350×10-6CO2处理的植株分别下降11%和提高25%。
2.4 干物质分配
本研究表明,根际NaCl胁迫和大气CO2富集都显著影响植物干物质分配及在植株不同器官中的分布比例(图1)。试验结果证明,根际NaCl盐渍抑制干物质在叶片中积累和向果实中分配,大气CO2富集则促进植物干物质向旺盛生长的器官中分配,这种趋势在叶片和果实中尤其明显。本试验处理组合中,干物质向主干中的分配不如其他器官明显(尤其是与叶片和果实相比)。这进一步证实,在本试验期间,供试植株的叶片和果实分别是营养生长和生殖生长的主导中心,而主干则是次要的生理活动器官。所有处理中,经根际25 mg·L-1NaCl胁迫和大气350×10-6CO2处理的植株形成干物质最少;而根际无NaCl盐渍、大气CO2为1 050×10-6处理的植株合成干物质最多(图1)。
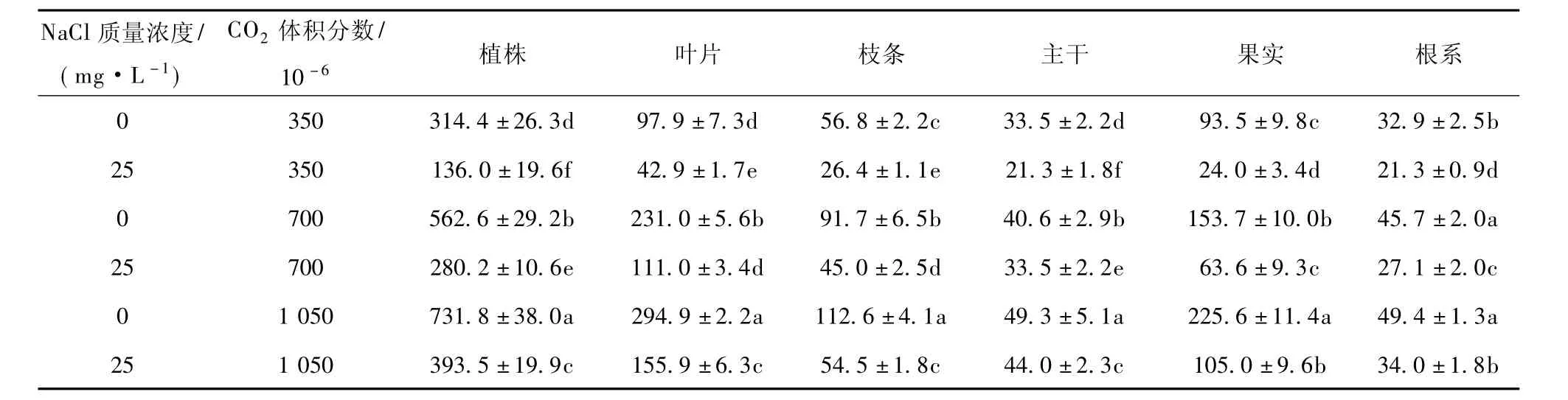
表3 根际NaCl盐渍和大气CO2富集对人参果植株和器官干物质日增长量(mg·株-1·d-1)的影响Table 3 Daily increment of dry mass(mg·plant-1·d-1)in pepino plants and organs as affected by NaCl salinity and CO2enrichment

图1 根际NaCl盐渍和大气CO2富集对人参果植株干物质向不同器官相对分配比例的影响(同一柱形右侧的不同字母表示5次重复中置信度为95%的显著性差异)Fig.1 Relative distribution of dry mass in different organs of pepino plants as affected by NaCl salinity and CO2enrichment(Different letters on the right side of the same column denote significant differences at P<0.05,n=5)
从总体上看,干物质在植株不同器官中的相对分布,以叶片最多、果实其次、根系再次、枝条较少、主干最少。这些事实再次证明,植物叶片和果实比其他器官对大气CO2富集的反应更敏感。植物干物质分配到叶片和果实中的比例虽因根际NaCl胁迫而显著下降,但随着大气中CO2的升高而增加。相反,植株干物质向主干和根系中的分配,却因根际盐渍胁迫而增加,并随大气CO2的富集而减少。这些现象主要是由于植物适应自然的生理代谢趋势和体内干物质分配的生物自动调节机理所致。但是,干物质在枝条中的分配,无论是受根际NaCl盐渍还是受大气CO2富集的影响,反应都不明显。这主要是因为该器官在供试期间处于生理代谢的迟缓时期所致(图1)。
2.5 植株耗水量
根际盐渍处理,不仅明显减少植株耗水量,而且显著降低根系吸收水分的效率和植株水分利用效率(表4)。大气CO2富集显著增加植株的耗水量和水分利用效率,但并未明显提高植物根系的吸水效率(表4)。这些现象表明升高大气CO2对植物根系生理功能指标(如根系吸水效率和植株水分利用率)的影响比其他与植物耗水有关的生理代谢指标(如植物干物质生产效率和光合作用水分利用效率;Chen et al.,1999b)要小。因而,经25 mg·L-1NaCl盐渍处理植株的耗水量和水分利用率,比在350×10-6CO2条件下未经盐渍处理者,分别下降31%和37%;当大气CO2提高到700×10-6及1 050×10-6时,这两项指标相应提高11%和61%及17%和99%。由于根际NaCl胁迫和大气CO2富集的互作关系,经25 mg·L-1NaCl盐渍和750×10-6及1 050×10-6CO2处理的植株,依次比对照的耗水量减少25%及18%,而相同植株的水分利用率得以提高。这说明大气CO2富集能使植物干物质生产过程中更有效并高效地利用水分。
2.6 叶片蒸腾系数和植株日耗水量
本试验中,350×10-6CO2下经25 mg·L-1NaCl处理的植株叶片蒸腾系数最大,而在1 050×10-6CO2下根际无NaCl胁迫的植株叶片蒸腾系数最小(图2)。相比之下,供试植株的叶片蒸腾系数在25 mg·L-1NaCl盐渍处理的单项因素影响下增加了56%,但当大气CO2体积分数从350×10-6提高到700×10-6和1 050×10-6时,该指标依次下降39%和58%。其中,受根际NaCl胁迫和大气CO2富集至700×10-6或1 050×10-6的互作效应影响,供试植株的叶片蒸腾系数下降4%~23%。这进一步证明,未来的全球大气CO2富集将有利于提高该作物的水分利用效率。

表4 根际NaCl盐渍和大气CO2富集对人参果植株含水量和耗水量的影响Table 4 Water content and water consumption of pepino plants as affected by NaCl salinity and CO2enrichment
另一方面,本研究还观察到植株日耗水量受根际NaCl胁迫而减少,但随大气CO2升高而增加(图2)。这说明根际NaCl盐渍对供试植株日耗水量的影响比大气CO2富集的效应更显著,主要是因为前者比后者不仅对植物根系吸水有更直接的抑制作用,而且对植株的需水和用水也有更显著的影响。在大气350×10-6CO2的情况下,25 mg·L-1NaCl盐渍处理平均减少供试植株31%的日耗水量。当大气CO2体积分数升高1倍和2倍时,无NaCl胁迫的植株日耗水量分别增加11%和17%,而受NaCl盐渍的植株日耗水量相应提高25%和18%。这些事实证明,在根际受到NaCl盐渍的条件下,提高大气CO2能有效地调节该作物的耗水量。
3 讨论
3.1 大气CO2富集和根际NaCl盐渍的作用及其互作效应
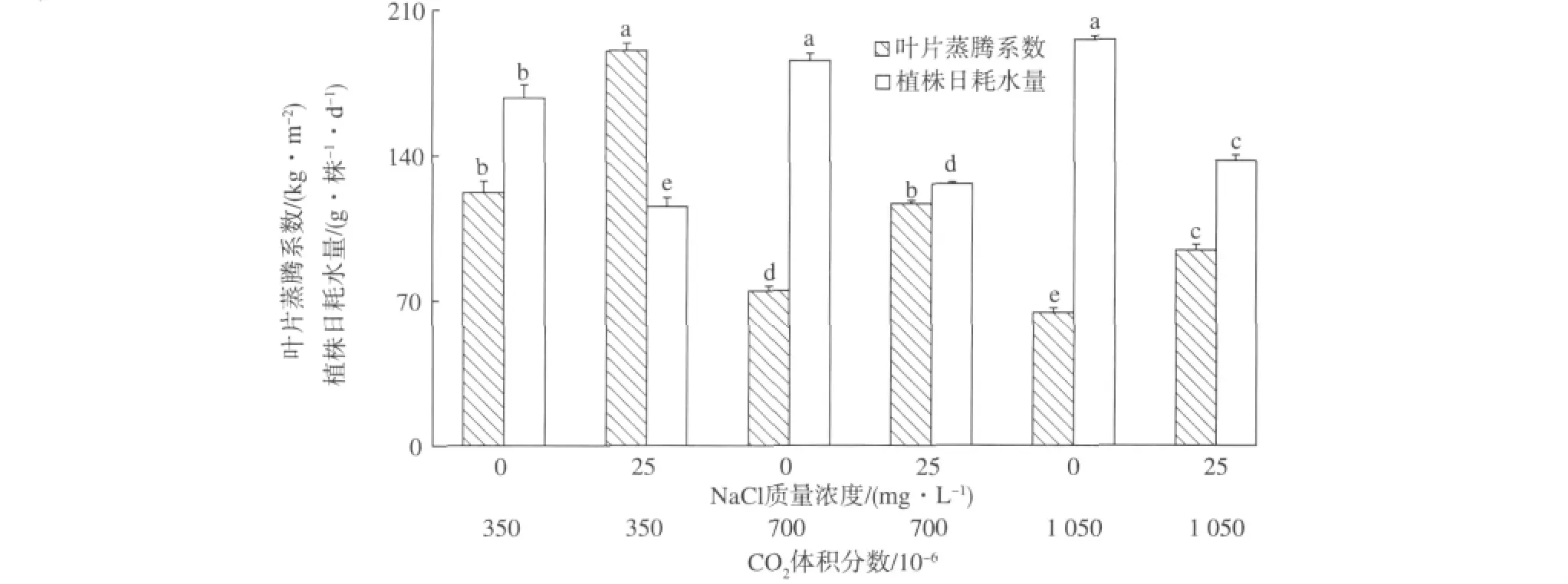
图2 根际NaCl盐渍和大气CO2富集对人参果叶片蒸腾系数和植株日耗水量的影响(同一柱形顶部的不同字母表示5次重复中置信度为95%的显著性差异)Fig.2 Leaf transpiration index and daily water consumption of pepino plants as affected by NaCl salinity and CO2enrichment(Different letters on the top of the same column denote significant differences at P<0.05,n=5)
在20世纪,全球大气平均CO2体积分数从270×10-6上升至350×10-6以上。这种趋势现在还在继续,并以每年1.5×10-6~2.0×10-6的速率上升。预计到21世纪末全球大气CO2还将翻番(Bazzaz,1990)。大气CO2的升高会通过促进植物的生长发育而增加作物生产力(Eamus and Jarvis,1989)。Kimball(1983)统计分析了先前对37种作物的430个试验结果,认为如果大气CO2在目前的水平上翻番,它们的产量将平均提高33%±6%。这种效应直接来自于升高大气中CO2,从而促进作物光合作用同化能力和干物质合成水平。Manderscheid and Weigel(1995)认为,近代德国作物产量的提高主要是直接得益于大气CO2的富集作用。若全球大气CO2以每年0.5%~0.7%的速度上升,当大气CO2每上升1×10-6,该国的作物平均产量即可提高0.25%~0.47%。研究发现,当大气CO2体积分数从350×10-6升高到700×10-6和1 050×10-6时,无NaCl盐渍及受25 mg·L-1NaCl盐渍的人参果经济产量分别提高64.73%和133.64%及163.39%和329.83%,折合成每升高1×10-6CO2对提高其产量的效应依次为0.18%和0.19%及0.46%和0.47%(Chen et al.,1999a)。本研究结果不仅与上述事实相符,而且进一步提供了大气CO2富集能促进植物干物质生产和提高水分利用率的科学证据。另外,全球大气中CO2升高还会导致空气温度的上升和降雨模式及数量的改变,这些变化对植物的光合作用、水分利用效率和干物质生产都会产生较大影响(Pandey et al.,2007;Thongbai et al.,2010)。因此,今后还有待更系统深入的研究,才能更全面和客观地评估其对作物生产的潜在效应。
农田中NaCl盐渍包括土壤和灌溉水中存在NaCl成分。这种情况会显著影响作物营养生长和生殖生长(Chen et al.,1999a)、光合作用能力(Chen et al.,1999b)和干物质生产(表2),以及植株器官中干物质的增长(表3)、积累和分配(图1),及其利用水分的效率(表4)。迄今为止,全球盐渍化土壤的总面积已达4×108~9×108hm2,造成70%的作物生产力下降(Lenssen et al.,1993)。因此,通过有效地解决这个问题来进一步提高作物生产力是在今后不断变化的全球气候和环境条件下很重要的研究课题。
由于作物内在的遗传特性,不同种类的作物对根际NaCl盐渍和大气CO2富集的表现存在差异(Idso and Idso,1994;Tester and Langridge,2010),从而对正确预测在大气—植物—土壤—水分的农田生态系统中,全球大气中CO2升高和农田土壤中NaCl胁迫对作物生产及其水分利用的复合互作效应带来更大的困难。因为作物在光合作用和呼吸作用过程中通过叶片气孔使其叶片和大气进行水汽和CO2分子的交换,而其植株主要通过根系从土壤中吸收水分,再经过主干和枝条把水分输送到叶片和果实等生理代谢中心,最后通过叶片气孔把水分蒸腾到大气中。对植株来说,上述一系列的生理过程会进一步制约其光合生产能力,叶片和根系生长发育及其他生理生化代谢,从而形成对外界环境的变化进行自我调节和适应等生物反应(Lenssen et al.,1993;Rogers and Runion,1994)。据Melgar et al.(2008)报道,大气中CO2升高能增强油橄榄树对土壤盐渍的适应能力。Martínez-Ballesta et al.(2004)也注意到,在土壤NaCl盐渍等环境胁迫的情况下,辣椒植株能对其不同的器官进行水分代谢和气体交换自我调节,本研究在人参果植株上观察到同样的现象和变化趋势。
此外,先前对CO2富集的许多研究结果都基于充足的养分供应条件(Bazzaz,1990)。然而,在大多数农田中经常缺乏足够作物生长所需的营养。这种情况在未来全球大气CO2进一步升高的条件下将更普遍。因为一方面大气CO2富集会有利于作物的碳素同化作用,但另一方面它会导致作物必需营养成分的失衡,尤其是维持植株碳氮供应比例的协调(Makino et al.,1997;Reich,2009;Bloom et al.,2010)。García-Sánchez and Syvertsen(2006)也观察到,在植物生长季节中提高大气CO2能增强Cleopatra柑桔和Carrizo枳橙实生苗耐盐渍的能力。Tattini and Traversi(2009)还发现,植物根际Ca2+浓度的升高有助于增强油橄榄树对土壤NaCl盐渍的生理抵抗力。另外,由于植物根际NaCl积累造成的水分逆境会进一步影响其根系吸收和利用营养的能力。本试验还观察到,根际受到25 mg·L-1NaCl盐渍的供试人参果植株不仅生长发育受阻,而且进一步出现营养代谢不良的症状。因此,根据上述科学假设和客观事实,今后有必要通过研究根际NaCl盐渍和大气CO2富集对植物营养吸收和利用的影响,更深入地探讨它们之间的复合互作关系。
3.2 干物质生产和分配
人参果植株干物质生产及其向不同器官中分配比例的改变,是对根际NaCl盐渍和大气CO2富集的重要生理反应和生态适应表现。本研究不仅证实了该植物对上述环境变化的反应与Lenssen et al.(1993)和Rozema et al.(1991)对其他C3植物的试验结果类似,而且还观察到供试植物的干物质生产效率(表2)及其向不同器官中的分配因被根际NaCl胁迫而显著受阻,但又因大气CO2升高而显著提高。本试验结果表明,植株干物质增长量因根际存在25 mg·L-1NaCl的胁迫而平均减少50%~54%,同时又因大气CO2体积分数从350×10-6升高到700×10-6和1 050×10-6而增加79%~189%(表3)。另外,本研究观察到的根际NaCl盐渍和大气CO2富集对人参果植株干物质生产的作用与对该作物叶片光合作用的影响(Chen et al.,1999b)相符。因此,根据上述结果可以推测,将来全球大气CO2进一步升高将能提高作物干物质生产能力,特别是在土壤盐渍化的农产区及由于NaCl积累而使灌溉水质退化的农田,会更有益处。
在本试验中,根际NaCl盐渍不仅会严重影响该作物叶片生长发育(表1),而且还会显著限制植株干物质生产力(表2)及其日增长量(表3)和分配到植株各器官中的数量(图1)。而且这些影响在叶片和果实中尤为明显,因为它们是植株生理代谢最旺盛的器官,所以对不利环境条件胁迫的反应最敏感。Matar et al.(1975)也观察到菠菜和生菜因根际受到NaCl胁迫而致使植株干物质生产能力下降。这些事实证明,根际NaCl胁迫对作物干物质生产力的负面影响,是农业生产中普遍存在的问题,在将来发展农业生产中必须高度重视,尤其是在盐碱土壤地区和沿海沼泽地区。此外,本研究发现根际NaCl胁迫同时限制干物质向地上部和地下部器官中分配(图1)。本试验中受NaCl盐渍的植株比未受NaCl胁迫者显著减小,这主要是由于盐渍逆境显著减少了植株干物质生产和分配(表3)。相反,大气中CO2富集能促进植物干物质生产(表2)、积累(表3)和分配(图1),主要是因为CO2是植物进行光合作用、生产干物质的必需原料。由此可以预测,由于未来大气CO2富集而引起的全球气候变化和能添加CO2的保护农业设施(如能控制微环境的温室和塑料大棚),都将有利于增强植物对根际土壤NaCl胁迫的抵抗力(Rogers and Runion,1994)。本研究还支持了Rozema et al.(1991)及Zeroni and Gale(1989)对耐盐植物Scirpus maritimus和Puccinellia maritime及玫瑰栽培品种“Sonia”受根际NaCl胁迫和大气CO2富集互作反应的试验结果,因而具有很好的代表性和广泛的适用性。
本研究进一步发现,升高大气CO2会增加干物质向植物地上部器官中分配,同时降低干物质向植物根系中分配的比例(图1)。而根际NaCl盐渍能促进干物质向植物地下部器官中分配,减少其向叶片、枝条、主干和果实中分配(图1)。这些事实与Flore and Lakso(1989)的报道相符,同时说明植物能对外界环境变化产生应变能力。这种应变不仅能通过改变其干物质生产的库源关系来实现,而且还能通过其自身的调节去适应外界环境变化,以防止或减轻植株可能发生的生理生化伤害。根据本研究结果,进一步总结出植物对环境变化生理反应的机理包括:第一、增加干物质向生理活跃的器官(库)中分配,以优先保证其生长中心的生理代谢优势;第二、减少干物质向生理不活跃的器官(源)中分配,以节约光合产物,适应环境逆变。本研究同时支持Eamus and Jarvis(1989)关于植物体内干物质分配取决于其内部生理因素和外部环境条件对植株地上部营养生长和生殖生长的中心器官(叶片与果实)光合产物的合成与供应,以及地下部器官(根系)吸收和利用营养与水分平衡的科学机理假设,为今后提高该作物生产水平积累了有用的实验资料和提供了可靠的科学依据。
3.3 植物耗水和水分利用效率
植物吸收和利用水分是其另一个重要的生理代谢过程。这个过程直接受根际NaCl胁迫的制约,同时也间接被大气CO2体积分数变化而影响。本研究观察到,根际NaCl盐渍和大气CO2富集都会减少植株含水量,但这种变化均未达到显著差异水平(表4)。这是因为植株含水量是供试植物固有的遗传特性,而这种特性不易因外界环境的变化而立即显著改变。Chen et al.(1999b)发现,植物叶片的蒸腾作用和气孔导度都随大气CO2的升高和根际NaCl盐渍的加重而下降。本研究进一步证明植物水分生理代谢胁迫与根际NaCl盐渍有密切关系,后者是前者的重要诱因。而且,根际NaCl胁迫所诱导的水分生理逆境现象,可以用Chen et al.(1999a)观察到的由于盐渍造成植物营养生长和生殖生长受抑的机理来解释。值得注意的是,本研究进一步发现根际NaCl盐渍对植物叶片生长和发育的抑制作用机理(表1),表现在:一方面减少植株的叶片数量(Chen et al.,1999a),另一方面增加比叶干重(即提高叶片质量)。由于叶片是植物进行干物质生产的主要场所,因而这种关系进一步影响植株干物质生产能力(表2)。本研究结果还证明,根际NaCl盐渍限制植物光合作用进行干物质生产的源泉,即减少植株总叶面积及单株叶面积的增长(表1)和植物干物质的日增长量(表2)及其向各种器官中分配的比例(表3)。此外,供试植株还表现出干物质生产库容的减小,从而限制植株营养器官和生殖器官中干物质日增长量的下降(表3),同时减少光合产物在叶片中积累和向果实中分配(图1)。因此,被根际NaCl盐渍的植株生长受阻,干物质日增长量下降(表3),根/冠比值增大(表2),进而降低其根系吸收水分的效率和植株水分利用效率(表4)。
大气CO2富集对上述植物生理代谢则有相反的作用。升高大气CO2,不仅会促进植物生长发育和干物质生产(表2),而且能降低植物叶片的蒸腾系数(图2),从而改善植株水分利用效率(表4)。本试验结果与Chen et al.(1999b)观察到的大气CO2富集能通过促进叶片同化CO2和降低叶片蒸腾速率来提高植物光合作用水分利用效率的事实相吻合。本研究还进一步证实,在700×10-6和1 050×10-6CO2条件下,减小的叶片气孔导度(Chen et al.,1999b)部分地抵消了因升高大气CO2而增强植物干物质生产,从而增加的植株耗水量。植物叶片蒸腾系数代表植株单位叶面积的耗水能力。供试植株的叶片蒸腾系数因根际NaCl盐渍而提高,又随大气CO2升高而下降(图2)。这一方面与这些处理影响植物叶片的生长发育相联系(Chen et al.,1997a),另一方面与它们能改变叶片气孔开闭的程度有关(Chen et al.,1999b)。尤其是后者,会直接制约叶片的蒸腾作用能力。据Eamus and Jarvis(1989)报道,植物水分利用效率随大气CO2升高而显著提高。本研究进一步揭示,植物总耗水量(表4)并不与其植株个体发育大小和器官中干物质增长数量成相同的比例增长(表3),而植株水分利用效率的显著提高主要得益于在较高的大气CO2条件下叶片生理性能和自我调节功能的改善,但植物根系吸收水分的效率几乎无变化,这说明根系的生理功能较小地受到大气CO2富集的影响(表4)。上述因素的互作效应及其植物生理机制可以总结为:大气CO2富集能够通过抵消因盐渍而引起的植物生理干旱,改善根际NaCl胁迫对植株水分利用的负面效应。这是因为,首先,在较高的大气CO2下植物叶片有较强的光合作用能力,从而维持有效的干物质生产效率;其次,在大气CO2富集的情况下,植物能把叶片光合作用产物较多地输到根系,因而补偿盐渍逆境对根部生长的抑制;再次,在高CO2条件下,植物叶片气孔导度减小,能防止水气从叶片气孔向大气中散发,从而减小单位叶面积蒸腾速率;最后,由于植物蒸腾速率下降,其根系吸收水分和Na+及Cl-的能力也随之减弱(Rogers and Runion,1994),从而缓解根际受NaCl盐渍的胁迫。由于本研究结果仅在一次试验中获得,对于这些错综复杂的交互作用机理、过程和影响大小,还有待今后进一步开展系统的科学实验和深入探索。
4 结论
根际NaCl盐渍显著限制人参果植株干物质生产和水分吸收及利用。NaCl胁迫不仅阻碍叶片生长发育及干物质生产、积累和向植株其他器官分配,而且降低根系吸收水分效率和植株水分利用率(表5)。相反,大气CO2富集显著促进植物叶片发育、干物质生产及其向地上部和地下部器官分配。后者在植株生理代谢活跃的器官中表现更明显。升高大气CO2虽会增加植株总耗水量,但同时提高其水分利用效率(表5)。根际NaCl盐渍和大气CO2富集都显著影响植物叶片数量和质量。由于叶片是生理代谢中心,这会深刻影响植株干物质生产和水分利用,从而进一步影响其不同器官中干物质的增长和积累,及水分利用效率。因为上述因素的复合互作效应,将来全球大气CO2富集会有利于植物干物质生产和水分利用,并能抵消或补偿因根际受NaCl盐渍所产生的抑制作用(表5)。大气CO2富集与根际NaCl盐渍对比叶干重有叠加效应,而对其他的植物生理和形态指标则有抵消作用(表5)。因此,在有保护设施的农业生产中,适当升高其微环境中的大气CO2体积分数能促进作物干物质生产和分配,提高植株水分利用效率,即使在根际NaCl盐渍的条件下,也会有良好效果。这项技术适用于将来在塑料大棚和温室栽培短生长期作物时推广。对于这些因素的交互作用机理和有效调控措施,尚有待进一步深入研究和系统探索。

表5 根际NaCl盐渍和大气CO2富集对人参果植株干物质生产和水分利用的互作效应总结Table 5 A summary for the effects of NaCl salinity and CO2enrichment on dry mass production and water consumption of pepino plants
致谢:感谢UNTIEDT Ralf、LIBERT Helmut和BERG Harry先生,以及SARTER Sigrid女士对该试验和研究工作的技术帮助和支持。本文特为Prof.Dr.LENZ Fritz 80寿辰和Prof.Dr.JANSSENS Marc光荣退休而作。
Bazzaz F A.1990.The response of natural ecosystems to the rising global CO2levels[J].Ann Rev Ecol Syst,21:167-196.
Bloom A J,Burger M,Asensio J S R,et al.2010.Carbon dioxide enrichment inhibits nitrate assimilation in wheat and Arabidopsis[J].Science,328(5980):899-903.
Chen K,Hu G Q,Keutgen N,et al.1999a.Effects of NaCl salinity and CO2enrichment on pepino(Solanum muricatum Ait.)I.Growth and yield[J].Sci Hort,81:25-41.
Chen K,Hu G Q,Keutgen N,et al.1999b.Effects of NaCl salinity and CO2enrichment on pepino(Solanum muricatum Ait.)II.Gas exchange of leaves[J].Sci Hort,81:43-56.
Eamus D,Jarvis P G.1989.The direct effects of increase in global atmospheric CO2concentration on natural and commercial temperate trees and forests[J].Advances in Ecological Research,19:1-55.
Flore J A,Lakso A N.1989.Environmental and physiological regulation of photosynthesis in fruit crops[J].Hort Rev,11:111-157.
García-Sánchez F,Syvertsen J P.2006.Salinity tolerance of Cleopatra mandarin and Carrizo citrange citrus rootstock seedlings is affected by CO2 enrichment during growth[J].J Amer Soc Hort Sci,131:24-31.
Grattan S.2002.Irrigation water salinity and crop production[M].Oakland:University of California-Davis Agriculture and Natural Resources Publication:1-9.
Hoagland D R,Arnon D L.1938.The water culture method for growing plants without soil[M].Berkeley:College of Natural Resources at Univ of California-Berkeley:1-39.
Idso K E,Idso S B.1994.Plant responses to atmospheric CO2enrichment in the face of environmental constrains:a review of the past 10 years’research[J].Agric and Forest Meteorol,69:153-203.
Kimball B A.1983.Carbon dioxide and agricultural yield:An assemblage and analysis of 430 prior observations[J].Agron J,75:779-788.
Lenssen G M,Lamers J,Stroetenga M,et al.1993.Interactive effects of atmospheric CO2enrichment,salinity and flooding on growth of C3(Elymus athericus)and C4(Spartina anglica)salt species[J].Vegetatio,104/105(1):379-388.
Makino A,Harada M,Sato T,et al.1997.Growth and N allocation in rice plants under CO2enrichment[J].Plant Physiol,115:199-203.
Manderscheid R,Weigel H J.1995.Do increasing atmospheric CO2concentrations contribute to yield increases of German crops[J].J Agronomy and Crop Science,175(2):73-82.
Martínez-Ballesta M C,Martinez V,Carvajal M.2004.Osmotic adjustment,water relations and gas exchange in pepper plants grown under NaCl or KCl[J].Environ Exp Bot,52(2):161-174.
Matar Y,Doering H W,Marschner H.1975.Effect of NaCl and Na2SO4on dry matter production,mineral content and organic compounds of spinach and lettuce[J].Z Pflanzenernaehr Bodenkd,3:295-307.
Melgar J C,Syvertsen J P,Garcia-Sanchez F.2008.Can elevated CO2improve salt tolerance in olive trees?[J].J Plant Physiol,165:631-640.
Pandey R,Chacko P M,Prasad K V.2007.Higher than optimum temperature under CO2enrichment influrnces stomata anatomical characters in rose(Rosa hybrida)[J].Scientia Hort,113(1):74-81.
Piao S,Ciais P,Huang Y.2010.The impacts of climate change on water resources and agriculture in China[J].Nature,467:43-51.
Reich P.2009.Elevated CO2reduces losses of plant diversity caused by nitrogen deposition[J].Science,326(5958):1399-1402.
Rogers H H,Runion G B.1994.Plant responses to atmospheric CO2enrichment with emphasis on roots and the rhizosphere[J].Environ Pollut,83:155-189.
Rozema J,Dorel F,Janissen R.1991.Effect of elevated atmospheric CO2on growth,photosynthesis and water relations of salt marsh grass species[J].Aquat Bot,39(1/2):45-55.
Tattini M,Traversi M L.2009.On the mechanism of salt tolerance in olive(Olea europaea L.)under low-or high-Ca2+supply[J].Environ Exp Bot,65:72-81.
Tester M,Langridge P.2010.Breeding technologies to increase crop production in a changing world[J].Science,327(5967):818-822.
Thongbai P,Kozai T,Ohyama K.2010.CO2and air circulation effects on photosynthesis and transpiration of tomato seedlings[J].Scientia Hort,126(3):338-344.
Zeroni M,Gale J.1989.Response of‘Sonia’roses to salinity at three levels of ambient CO2[J].J Hort Sci,64:503-511.
