El Tor霍乱弧菌双精氨酸转运系统功能单位的分析研究
2012-01-23朱召芹景怀琦陈海丽胡芸文阚飙
朱召芹,景怀琦,陈海丽,胡芸文,阚飙
1. 上海市(复旦大学附属)公共卫生临床中心,上海201508; 2. 中国疾病预防控制中心传染病预防控制所,传染病预防控制国家重点实验室,北京102206
目前,霍乱在世界范围内仍是严重的公共卫生问题。据世界卫生组织(World Health Organization,WHO)专家会议估计,全球每年约发生550万例霍乱,其中以亚洲、非洲和拉丁美洲流行较为严重,引起亚洲约10万例和非洲约2万例患者死亡。尽管人类对引起霍乱流行的霍乱弧菌认识已久,但对其致病性及毒力相关功能基因尚不完全清楚。
双精氨酸转运系统(twin-arginine translocation system,Tat系统)是1997年Settles等[1]发现的存在于原核生物中的蛋白质运输系统,负责运输完全折叠蛋白,并结合一定的氧化还原因子[2],其特点是蛋白N端信号肽存在一保守的双精氨酸结构域(S/T-R-R-x-F-L-K)[3]。目前,Tat系统已陆续在大肠埃希菌、幽门螺杆菌、耶尔森鼠疫杆菌、伤寒沙门菌、铜绿假单胞菌、结核分枝杆菌、霍乱弧菌、流感嗜血杆菌、葡萄球菌等细菌染色体中被发现[4-6]。Tat系统不但在细菌生理功能方面,而且在一些病原微生物的致病性中发挥作用,推测可能也是一种新的毒性决定因素[1]。大多数细菌Tat系统在叶绿体膜和细胞膜中发挥作用[7,8]。已进行深入研究的细菌以革兰阴性大肠埃希菌和革兰阳性枯草杆菌为典型代表。大肠埃希菌的Tat系统需3种不同功能的膜结合蛋白成分(TatA、TatB和TatC)。目前革兰阴性菌比较普遍的Tat系统模型是TatB和TatC构成复合体,识别并结合到双精氨酸信号肽。TatA形成寡聚体,在TatB和TatC协同作用下,很可能在蛋白转运过程中起通道作用。
氧化三甲胺(trimethylamine-N-oxide,TMAO)是存在于胞质并参与细菌呼吸能量代谢的重要物质,也是Tat系统的底物,广泛应用于细菌Tat系统的功能研究。在厌氧培养条件下,TMAO是细菌的唯一电子受体,如细菌能在TMAO培养基上生长[9],即说明TMAO可被Tat系统进行功能性转运,从而推测Tat系统具有功能[9,10]。
虽然我们的前期研究提示霍乱弧菌有功能性的Tat系统,且与霍乱弧菌生物膜的形成和毒力有一定的相关性,但目前对霍乱弧菌Tat系统基因簇的各基因功能尚不清楚,尤其是霍乱弧菌Tat系统基因簇横跨Ⅰ、Ⅱ号2个染色体,通过序列比对没有发现TatD基因存在。为深入研究霍乱弧菌Tat系统相关基因及功能,本研究通过构建系列缺失株、回补株,进一步分析其基因功能、基因簇各构成基因功能,并确定功能必需基因。
1 材料和方法
1.1 菌株及质粒
本研究所用菌株、质粒及其特征详见表1。
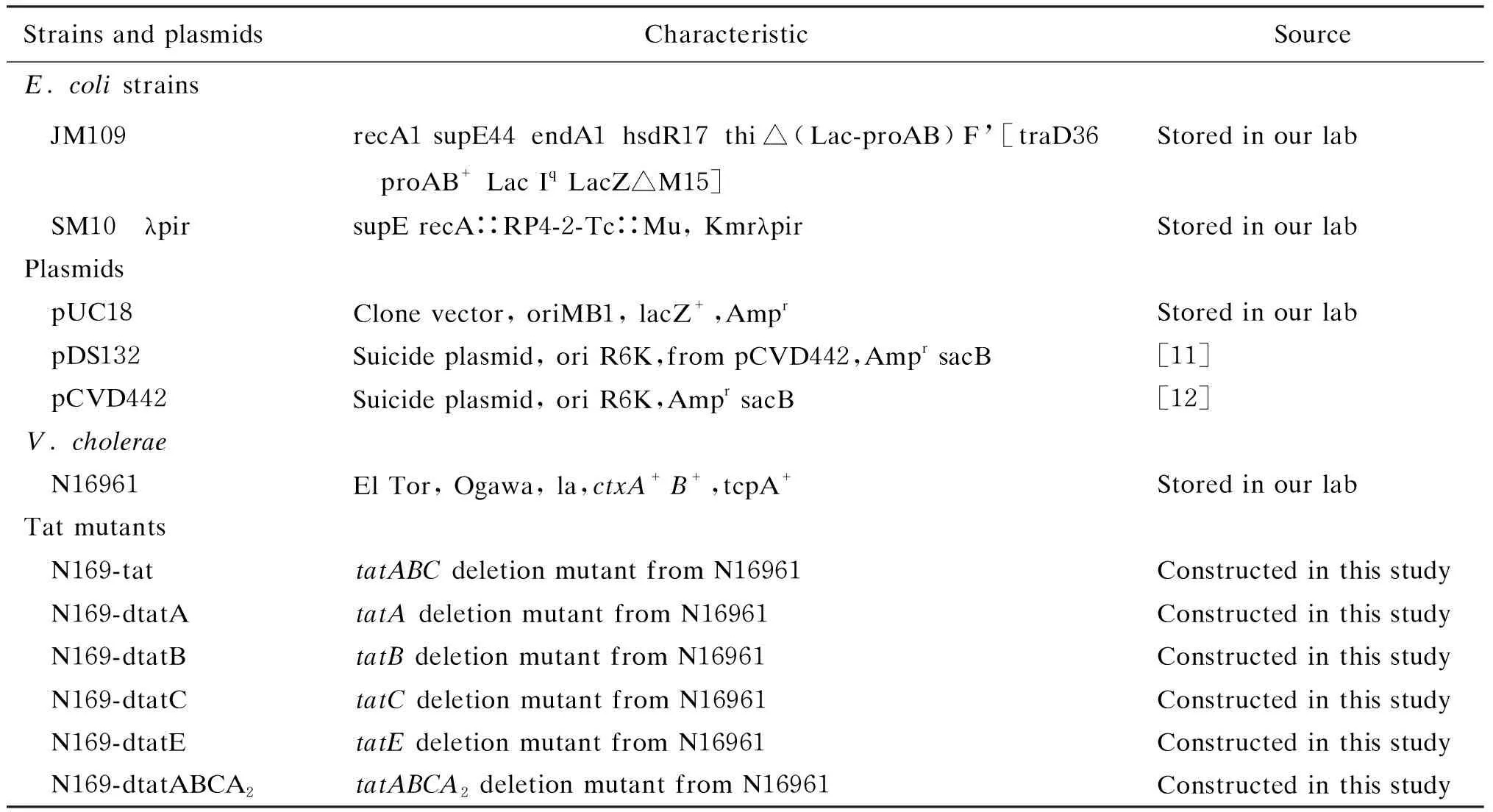
表1 研究所用的菌株和质粒Tab.1 The strains and plasmids used in the study
Tat系统基因缺失株的构建:将含有插入片段的pDS132[7]重组质粒与N16961中拟敲除片段的上下游序列同源重组,使部分基因或全部基因片段敲除。以含有插入片段pDS132重组质粒的SM10为供体菌,N16961野生株为受体菌,分别进行接合转移实验,获得接合子。接合子带有自杀质粒pDS132的氯霉素抗性和sacB基因(编码果聚糖蔗糖酶,对宿主菌在蔗糖浓度>5%的培养基上生长是致死性的)。首先用庆大霉素选择性培养基(或TCBS培养基)加氯霉素抗性平板进行阳性结合子筛选,然后用20%蔗糖培养基进行2次同源重组筛选,以排除目的基因只进行1次同源重组、自杀质粒仍未完全脱落的接合子,最后对筛选的缺失株进行鉴定。
1.2 试剂
TMAO、甲基紫精(methyl viologen,MV)、钼酸铵(ammonium molybdate)、硫胺素(thiamine,维生素B1)和亚硒酸钾(potassium selenite)购于Sigma公司,氯霉素购于上海生工生物工程技术服务有限公司。
1.3 周质和原生质体的分离
Tat系统可将转运底物从胞质至周质跨膜转运,因此测定细菌胞质和周质内转运底物的量可帮助判断Tat系统的功能。将霍乱弧菌N16961野生株和各缺失株用LB培养过夜,次日1∶100转种至新鲜培养基,待菌生长至OD600=0.8,离心收集细菌。按每1 g湿菌加入5 ml 0.2 mol/L pH 8.0 Tris缓冲液,将细菌沉淀物悬起,然后按每1 g湿菌加入5 ml 0.2 mol/L pH 8.0 Tris和34%蔗糖溶液。冰上加入0.5 mol/L乙二胺四乙酸(ethylenediaminetetraacetic acid,EDTA),使其终浓度达1 mmol/L并混匀。冰浴2.5 min。加入溶菌酶,使其终浓度为0.1 mg/ml,冰浴2.5 min。缓慢滴加预冷的三蒸水 10 ml,在5 min内滴完。13 400g离心20 min,上清液吸出,即为周质。沉淀物为原生质体。沉淀物用50 mmol/L Tris + 5 mmol/L MgCl2(pH 7.6)洗1次,超声破碎,13 400g离心,上清液为细胞质[2]。
1.4 非变性聚丙烯酰胺凝胶电泳钼酶活性的检测
基于对甲基紫可视化活性染色法,通过非变性聚丙烯酰胺凝胶电泳(native polyacrylamide gel electrophoresis,native-PAGE)检测霍乱弧菌野生株和缺失株细胞质和周质内TMAO还原酶的活性[4]。电泳结束前,将50 ml磷酸盐缓冲液放入500 ml量筒中,用一次性手套将量筒封好并用皮筋勒紧。缓慢从液体底部向缓冲液充氮气15 min,加入MV溶液500 μl,继续充氮气15 min。将非变性胶小心取下,浸泡在非变性电泳缓冲液中待用。将充完氮气的磷酸盐缓冲液快速倒出25 ml于一次性平皿中,加入少量用0.01 mol/L NaOH溶解的NaS2O4,直至液体全部变蓝。对凝胶进行染色3~5 min至整个胶面呈深蓝色,移至含有酶作用底物TMAO的染液内继续染色数分钟。将胶移至两胶片中以隔绝空气,通过酶与底物作用,产生新生氧气,从而使染料氧化变白,即在该酶泳动条带处呈现白色条带。用凝胶成像仪或照相机记录结果[2]。
1.5 Tat底物TMAO(torA基因编码)的检测
用M9-TMAO基础培养基来检测Tat野生株和缺失株的Tat系统功能是否存在[13]。在厌氧培养条件下,TMAO是M9-TMAO基础培养基中唯一能提供能量的营养成分,同时TMAO又是公认的Tat系统转运底物,所以细菌在厌氧培养条件下对TMAO的利用情况可直接反映Tat系统功能。将培养基分装后,待检测菌株置37 ℃厌氧静止培养。测定不同培养时间的菌液浊度(OD600),据此判断细菌生长情况。
1.6 生物信息学分析
采用BLASTn程序在GenBank中搜寻与已知大肠埃希菌MG1655(K12)的Tat系统基因簇(tatA、tatB、tatC、tatD和tatE)DNA序列具有同源性的霍乱弧菌N16961基因,再进一步用Bioedit软件进行相应序列的比对。
1.7 统计学处理
用GraphPad Prism 5.0软件进行统计分析。采用one-way ANOVA对各组进行单因素方差统计分析。以P<0.05为差异有统计学意义。
2 结果
2.1 霍乱弧菌Tat系统基因簇的生物信息学分析
不同细菌中Tat系统基因的结构和组成不同。本研究通过对已测序的霍乱弧菌标准株N16961基因组与大肠埃希菌tatA、tatB、tatC、tatD操纵子同源性分析,预测霍乱弧菌N16961存在Tat系统 的主要基因(表2)。tatA(VC0086)、tatB(VC0087)和tatC(VC0088)位于Ⅰ号染色体上,与大肠埃希菌不同的是,在霍乱弧菌N16961Ⅰ号染色体上没有发现与大肠埃希菌tatE同源性高的基因片段,与大肠埃希菌tatE基因同源性高的基因tatA2(VCA0533)位于霍乱弧菌小染色体上(Ⅱ号染色体);tatA、tatB、tatC基因簇附近没有tatD基因。尽管霍乱弧菌的tatA2基因与大肠埃希菌的tatA和tatE基因编码蛋白的氨基酸序列均具有一定同源性(分别为36.7%和38.2%),但由于大肠埃希菌tatA、tatB、tatC、tatD与tatE位于不同操纵子下游,因此根据霍乱弧菌tatA2位置初步推测其在Tat系统功能上可能相当于大肠埃希菌的tatE。霍乱弧菌Tat系统各基因编码蛋白与大肠埃希菌对应基因编码蛋白的氨基酸同源性为43.3%~65.7%。除Tat系统基因外,电子传递链和能量代谢相关基因、细胞色素c551过氧化物酶基因(VC0089)和辅酶Q蛋白合成Aarf基因(VC0085)分别位于tatA、tatB、tatC操纵子的下游和上游。
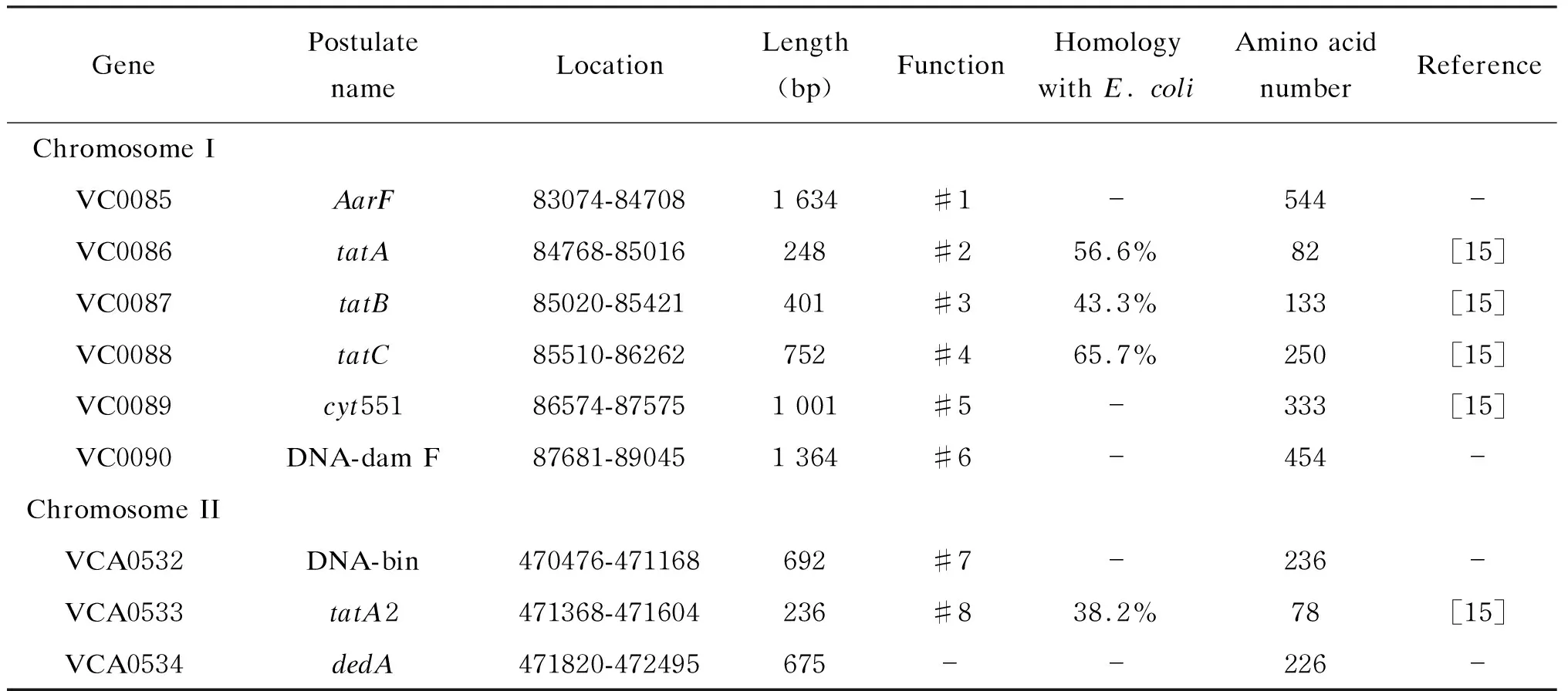
表2 N16961 Tat系统基因簇分析Tab.2 The analysis of gene cluster of Tat system in N16961*
2.2 霍乱弧菌Tat系统转运功能的研究
为确认经序列比较而推测的霍乱弧菌Tat系统是否具有蛋白运输功能,本研究比较霍乱弧菌的野生株与缺失株在厌氧培养条件下于TMAO基础培养基中的生长状况,观察细菌是否可利用TMAO作为唯一的能量来源。结果发现(图1、2),当霍乱弧菌N16961的Tat系统基因簇中tatA、tatB、tatC3个基因同时缺失或tatA、tatB、tatC和tatA24个基因同时缺失时,菌株不能在TMAO基础培养基中生长,提示这些基因的完整性对霍乱弧菌Tat系统功能很重要。单独缺失tatB或tatC基因时,细菌生长明显受限,说明这2个基因对Tat系统的功能很重要;而单独缺失tatA或tatA2基因时,细菌生长仅一定程度受限,提示两者存一时对Tat系统功能影响不大,但tatA和tatA2双基因同时缺失,Tat系统功能则完全受阻。
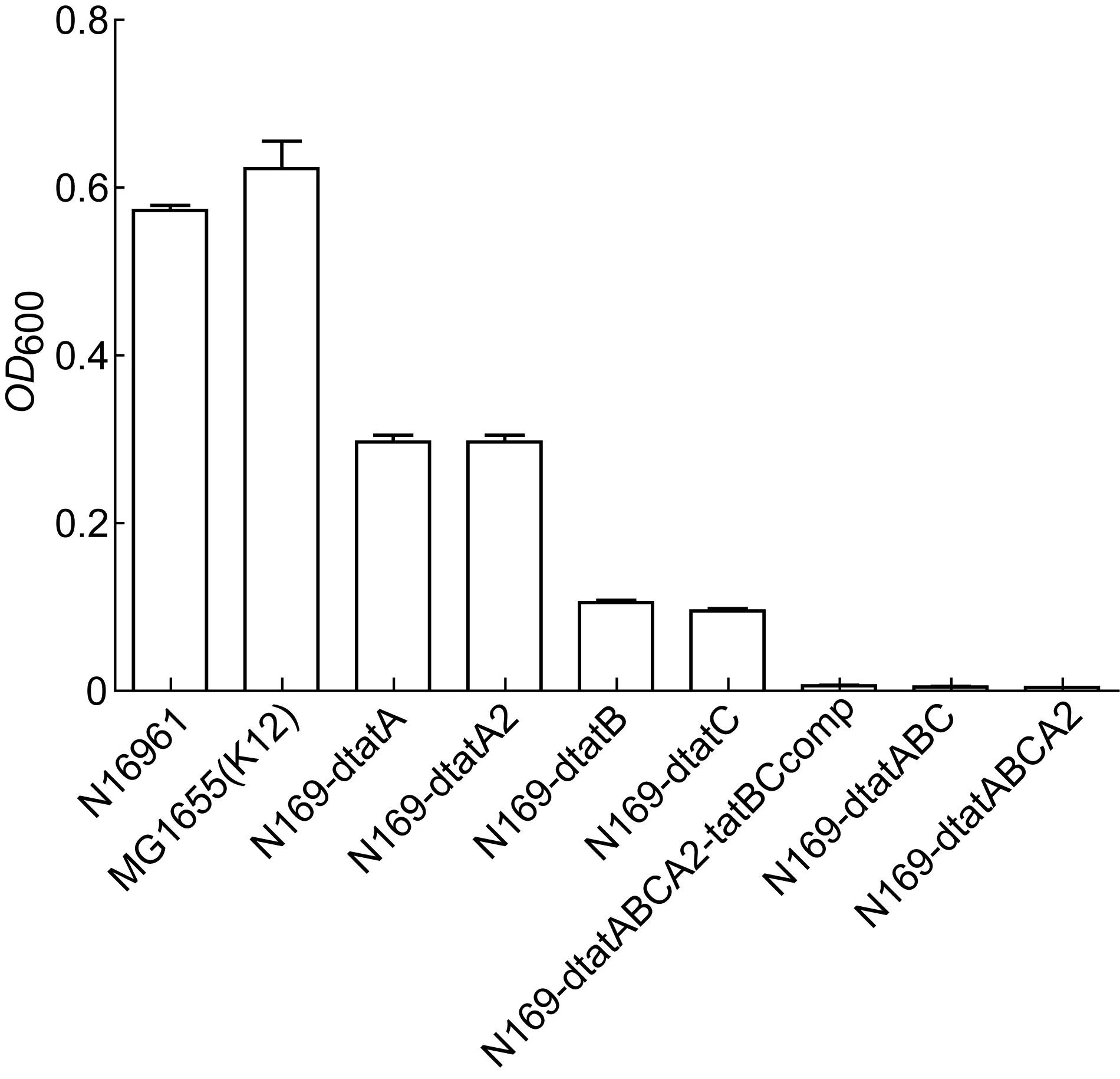
图1 厌氧培养条件下野生株和缺失株在M9-TMAO基础培养基的生长状况Fig.1 Wild type and mutant strains in M9-TMAO basal medium under anaerobic culture condition
2.3 霍乱弧菌Tat的转运底物
TMAO为Tat的转运底物,Tat可将TMAO从细菌胞质转运到周质,因此检测细菌胞质和周质TMAO活性成为细菌Tat功能验证研究中广泛使用的方法[4,13,14]。研究发现,霍乱弧菌N16961无论胞质还是周质中TMAO活性检测均为阳性,而Tat基因缺失株N169-dtatABC只在胞质样本检测到酶活性。为进一步确认霍乱弧菌存在功能性Tat分泌系统,本研究对Tat基因簇中很可能为Tat功能基因的缺失株和回补株进行检测,发现N169-dtatA(tatA单缺失株)、N169-dtatA2(tatA2单缺失株)及回补株N169-dtatABCA2-cp(pBAD24-tatA2BC)、N169-dtatABCA2-cp(pBAD24-tatABC)无论胞质还是周质酶活性检测均为阳性,提示tatA或tatA2单基因缺失不导致转运功能丧失;而缺失株N169-dtatABCA2、N169-dtatABC、N169-dtatBC和回补株N169-dtatABCA2-cp(pBAD24-tatBC)只在胞质样本检测到酶活性(图2),提示tatB和tatC是Tat系统功能必需基因。

C: Cytoplasm; P: Periplasm; 1: N169-dtatABCA2; 2: N169-dtatABC; 3: N169-dtatA; 4: N169-dtatA2; 5: N169-dtatABCA2-cp (pBAD24-tatBC); 6: N169-dtatBC; 7: N169-dtatABCA2-cp (pBAD24-tatA2BC); 8: N169-dtatABCA2-cp (pBAD24-tatABC).
3 讨论
近年来,对Tat系统的研究越来越受到重视。Tat系统是存在于病原菌中并有重要毒力机制的特殊蛋白转运系统,参与细菌胞膜形成、生物膜形成、电子运输链排列、多种环境压力下生存等。
霍乱弧菌Tat系统各基因编码蛋白与大肠埃希菌对应基因编码蛋白的氨基酸同源性为43.3%~65.7% ,但霍乱弧菌Tat系统基因簇与大肠埃希菌Tat系统存在很多差异。(1)在霍乱弧菌Ⅰ号染色体没有发现与大肠埃希菌tatE同源性高的基因片段;与大肠埃希菌tatE基因同源性高的基因tatA2(VCA0533)位于霍乱弧菌小染色体(Ⅱ号染色体)上,tatA2与大肠埃希菌tatA和tatE的同源性均很高。(2)霍乱弧菌Tat系统的主要基因位于霍乱弧菌的2个染色体上,Ⅰ号染色体tatA、tatB、tatC基因簇附近没有tatD基因。(3)除Tat系统基因外,电子传递链和能量代谢相关基因〔细胞色素c551过氧化物酶基因(VC0089)和辅酶Q蛋白合成Aarf基因(VC0085)〕分别位于tatA、tatB、tatC操纵子的下游和上游。因此,研究霍乱弧菌Tat系统各基因的功能对研究Tat系统对霍乱弧菌蛋白转运、致病力和环境生存作用是非常必要的。
本研究显示,tatA或tatA2单基因缺失均不影响Tat系统功能;tatA和tatA2同时缺失,Tat系统功能受到影响,提示霍乱弧菌中tatA或tatA2两者存一即可使Tat系统发挥功能,但不能同时缺少tatA和tatA2,显示这2个基因是Tat系统功能发挥所必需的。这与大肠埃希菌tatA与tatE的功能可互相替换[16]相似,同时根据大肠埃希菌tatA、tatB、tatC基因簇与tatC基因的位置关系,初步判定霍乱弧菌的tatA2功能更接近大肠埃希菌的tatE。
tatB和tatC单基因缺失可导致霍乱弧菌Tat系统功能严重受损,因此初步判定它们是霍乱弧菌Tat系统的必需基因。这也与霍乱弧菌的tatB和tatC基因功能相似。分析大肠埃希菌Tat系统各组分的稳定性,发现当tatB缺失后,tatC会很快降解,所以tatB对维持tatC的稳定性发挥重要作用[16]。霍乱弧菌tatB单基因缺失引起Tat系统功能受损是tatB缺失直接导致的,还是因为其影响tatC的稳定性间接导致的,需进一步研究。
本研究通过生物信息学分析和功能验证实验,推测了霍乱弧菌编码Tat系统的基因簇,验证了这些基因的生物学功能。结果提示,tatB和tatC是霍乱弧菌Tat系统功能必需的,tatA与tatA2基因功能重复,但整个Tat系统需要tatA或tatA2,两者具备一个即可。由此推断tatA-tatB-tatC或tatA2-tatB-tatC是霍乱弧菌Tat系统功能最小单位,为进一步研究霍乱弧菌毒力及致病性相关功能基因提供了重要理论依据。
[1] Settles AM, Yonetani A, Baron A, Bush DR, Cline K, Martienssen R. Sec-independent protein translocation by the maize Hcf 106 protein [J]. Science, 1997, 278(5342): 1467-1470.
[2] Santini CL, Ize B, Chanal A, Müller M, Giordano G, Wu LF. A novel sec-independent periplasmic protein translocation pathway in Escherichia coli [J]. EMBO J, 1998, 17(1): 101-112.
[3] Berks BC, Sargent F, Palmer T. The Tat protein exports pathway [J]. Mol Microbiol, 2000, 35 (2): 260-274.
[4] Pradel N, Ye C, Livrelli V, Xu J, Joly B, Wu LF. Contribution of the twin arginine translocation system to the virulence of enterohemorrhagic Escherichia coli O157∶H7 [J]. Infect Immun, 2003, 71(9): 4908-4916.
[5] Ding Z, Christie PJ. Agrobacterium tumefaciens twin-arginine dependent translocation is important for virulence,flagellation,and chemotaxis but not type Ⅳ secretion [J]. J Bacteriol, 2003, 185(3): 760-771.
[6] 张丽娟, 高守一, 景怀琦, 阚飙, 祁国明, 刘延清, 吴龙飞, 徐建国. 霍乱弧菌Tat 蛋白运输系统基因簇的确定与功能阻断分析[J].遗传学报, 2002, 29(10): 936-940.
[7] Robinson C, Bolhuis A. Tat-dependent protein targeting in prokaryotes and chloroplasts [J]. Biochim Biophys Acta, 2004, 1694(1-3):135-147.
[8] Müller M, Klösgen RB. The Tat pathway in bacteria and chloroplasts (review) [J]. Mol Membr Biol, 2005, 22(1-2): 113-121.
[9] Bogsch EG, Sangent F, Stanley NR, Berks BC, Robinson C, Palmer T. An essential component of a novel bacterial protein export system with homologues in plastids and mitochondria [J]. J Biol Chem, 1998, 273(29): 18003-18006.
[10] Stanley NR, Findlay K, Berks BC, Palmer T. Escherichia coli strains blocked in Tat-dependent protein export exhibit pleiotropic defects in the cell envelope [J]. J Bacteriol, 2001, 183(1): 139-144.
[11] Philippe N, Alcaraz JP, Coursange E, Geiselmann J, Schneidera D. Improvement of pCVD442, a suicide plasmid for gene allele exchange in bacteria [J]. Plasmid, 2004, 51(3): 246-255.
[12] Donnenberg MS, Kaper JB. Construction of an eae deletion mutant of enteropathogenic Escherichia coli by using a positive-selection suicide vector [J]. Infect Immun, 1991, 59(12): 4310-4317.
[13] Xiong Y, Santini CL, Kan B, Xu J, Filloux A, Wu LF. Expression level of heterologous tat genes is crucial for in vivo reconstitution of a functional Tat translocase in Escherichia coli [J]. Biochimie, 2007, 89(5):676-685.
[14] Kikuchi Y, Date M, Itaya H, Matsui K, Wu LF. Functional analysis of the twin-arginine translocation pathway in Corynebacterium glutamicum ATCC 13869 [J]. Appl Environ Microbiol, 2006, 72(11):7183-7192.
[15] Zhang L, Zhu Z, Jing H, Zhang J, Xiong Y, Yan M, Gao S, Wu LF, Xu J, Kan B. Pleiotropic effects of the twin-arginine translocation system on biofilm formation, colonization, and virulence in Vibrio cholera [J]. BMC Microbiol, 2009, 9:114.
[16] Sargent F, Stanley NR, Berks BC, Palmer T. Sec-independent protein translocation in Escherichia coli. A distinct and pivotal role for the TatB protein [J]. J Biol Chem, 1999, 274(51): 36073-36082.
