安徽皇藏峪自然保护区青檀林的群落特征
2012-01-04尤海梅于法展阎传海
尤海梅,于法展 ,阎传海
(1.江苏师范大学 城市与环境学院,江苏 徐州 221116;2.青岛理工大学 环境与市政工程学院,山东 青岛 266033)
安徽皇藏峪自然保护区青檀林的群落特征
尤海梅1,于法展1,阎传海2
(1.江苏师范大学 城市与环境学院,江苏 徐州 221116;2.青岛理工大学 环境与市政工程学院,山东 青岛 266033)
为了解皇藏峪自然保护区内天然青檀林的发展状况、有效地保护青檀群落,采用样方调查法,对青檀群落的种类组成、结构、外貌特征进行了调查研究。结果表明:群落中有维管束植物61种,隶属39科56属。植物区系中热带成分和温带成分均占有较大比例,但以温带属种居多,呈现出由温带向亚热带过渡的特点。群落的组成物种以落叶高位芽植物为主,地面芽植物也占有较大比例。群落垂直结构简单,可分为乔木层、灌木层、草本层,青檀占绝对优势,林下更新良好,优势种群多呈增长型,群落处于稳定的生长状态。同时,人为干扰(如砍割灌草等)造成的大量青檀萌生个体呈多株丛生状集中分布于灌草层,影响着林下植物的生长和群落的稳定,建议加强对林下植被的保护。
青檀;群落;物种组成;结构;外貌;皇藏峪自然保护区
青檀Pteroceltis tatarinowii是第三纪古热带植物区系的孑遗植物,也是我国特产的单种属植物[1-2];它集造纸、材用、药用、饲料、景观、生态防护和科学研究等多种价值于一体,是国家Ⅲ级珍稀濒危保护植物[3-4]。目前有关青檀的研究主要集中在青檀的形态解剖学、生理学、繁殖生物学、种群生态学、育苗及栽培技术、檀皮质量与加工等方面[5-13],而有关青檀群落生态学的研究报道较少[1-2]。青檀在我国分布很广,自华北、西北至华南、贵州、四川、西藏均有分布[14],但多是零星的散生,集中分布的较为稀少[1]。一些研究发现,现存较少的青檀林中青檀的天然更新状况大都不良,现有的青檀林多处于不稳定、难持续的状态[1,4,15]。因此,研究青檀林的群落学特征,对青檀树种的可持续利用与保护具有重要意义。同时,青檀也是我国长城以南石灰岩山地造林的先锋树种[1],对其群落进行研究,有助于石灰岩山地的植被恢复与重建。
安徽省皇藏峪自然保护区内保存有一定数量的块状集中分布的天然青檀林,是该区的特色森林类型之一[16]。但到目前为止,有关该区青檀林的详细研究尚仅涉及种群生态学[17]。研究该区青檀林的群落学特征,有助于深入了解该群落的种类组成与结构特征、更新特点及发展动态,可为有效地保护和构建青檀群落、持续地利用青檀资源提供科学参考。
1 研究区与研究方法
1.1 研究区概况
皇藏峪自然保护区位于安徽省萧县东南部,地 处 东 经 117°03´~ 117°06´、 北 纬 34°00´~34°06´之间。保护区的地貌类型为岛状残丘,系山东古老丘陵向南的延伸,属喀斯特石灰岩山地,最高峰海拔389 m。保护区地处暖温带季风气候区,年均温度14.3℃,极端最低气温-24℃,极端最高气温41℃,≥10℃的年均积温4 800℃,年均降水量850~900 mm,降雨多集中在7、8月,无霜期约210 d。主要土壤为褐色石灰土,呈中性至微碱性。林下地形主要为浅沟和山坡,浅沟中存有大量的冲积碎石,缓坡土层厚、裸露岩石少,陡坡土层薄、裸露岩石多。皇藏峪自然保护区的地带性植被为暖温带落叶阔叶林,由于受人为破坏较轻,区内的落叶阔叶林保存较为完好。
1.2 样地设置与群落调查
在皇藏峪自然保护区内的瑞云寺、仙人床附近,选取青檀占优势的林分设置3块20 m×20 m的调查样地,将每块样地划分为16个5 m×5 m的乔木样方,对乔木样方中高于或等于2.0 m的立木进行每木调查,记录种名、树高、胸径、冠幅、实生或萌生特性。在每个乔木样方内分别沿对角设置2个2 m×2 m的灌木样方和4个1 m×1 m的草本样方,调查灌木层和草本层的植物种类、数量、盖度、高度、实生或萌生特性等。同时记录样地的海拔、地形、坡向、坡位、坡度、土壤、岩石裸露度等环境因子,样地的基本情况见表1。

表1 调查群落基本情况Table 1 Basic characteristic of plots
1.3 数据处理
1.3.1 重要值的计算
采用重要值法分别乔木层、灌木层和草本层判定各物种在群落中的地位,计算公式为:
重要值=(相对密度+相对优势度+相对频度)/3式中,乔木层树种的相对优势度为树种的相对显著度,即该树种的胸高断面积占林分总胸高断面积的百分比;灌草层植物的相对优势度为物种的相对覆盖度。
1.3.2 径级结构的划分
根据树种的生活史特征和样地调查数据,以高度和胸径大小为标准进行立木径级的划分,其中树高小于或等于33 cm的树木划分为幼苗(即第1径级),树高大于33 cm的树木,按胸径每隔4 cm划分为一个等级,共划分13个径级,统计各径级树木的株数,分析种群的径级结构。
2 结果与分析
2.1 种类组成
皇藏峪3块青檀林样地中共出现维管束植物61种,隶属于39科56属。群落的物种组成虽不丰富,但科属组成却较为复杂。出现物种数最多的科是百合科Liliaceae(5属6种),其次是禾本科Gramineae(5属5种)和豆科Leguminosae(4属5种),呈单属种分布于青檀群落的科共30个(占总科数的76.9%)。呈单种分布于青檀林的属有51个(占总属数的91.1%),占绝对优势;出现2个物种的属有5个(朴树属Celtis、胡枝子属Lespedeza、土麦冬属Liriope、苔草属Carex和苦卖菜属Ixeris),占总属数的8.9%。青檀林中出现的61种植物均为被子植物,其中木本植物24科30属32种,以豆科(5属6种)和榆科Ulmaceae(2属3种)为主;草本植物16科26属29种,以禾本科(5属5种)、百合科(4属5种)、菊科Сompositae(3属4种)居多。
2.2 区系成分
按照吴征镒等[18-20]有关种子植物科、属分布区类型划分系统,将皇藏峪青檀林中出现的种子植物39个科划分为4种分布区类型,56个属划分为12种分布区类型(见表2)。在39科中,世界分布科共17科,占总科数43.6%,多为林下常见草本,如禾本科、菊科、豆科、莎草科Сyperaceae等;热带分布科共15科,占38.5%,且除热带亚洲和热带美洲间断分布的马鞭草科Verbenaceae外,其余均为泛热带分布科;温带分布科共7科,占17.9%,均为北温带分布科,如壳斗科Fagaceae、漆树科Anacardiaceae、胡颓子科Elaeagnaceae、山茱萸科Сornaceae等。在56属中,温带分布类型26属,占出现总属数的46.4%;热带分布类型22属,占总属数的37.5%,以泛热带成分为主;世界分布类型共7属,占总属数的12.5%;中国特有类型2属,占总属数的4.1%。可见,该区青檀林种子植物科的分布区类型以世界成分为主,热带和温带成分均占有较大比例,说明该区青檀群落的种类组成与热带、温带有着密切的联系,区系成分较为复杂;而植物属的分布类型以温带成分为主,说明该区种子植物区系具有明显的温带性质;热带成分的丰富存在,反映了该区青檀群落植物区系由温带区系向亚热带区系过渡的特点。
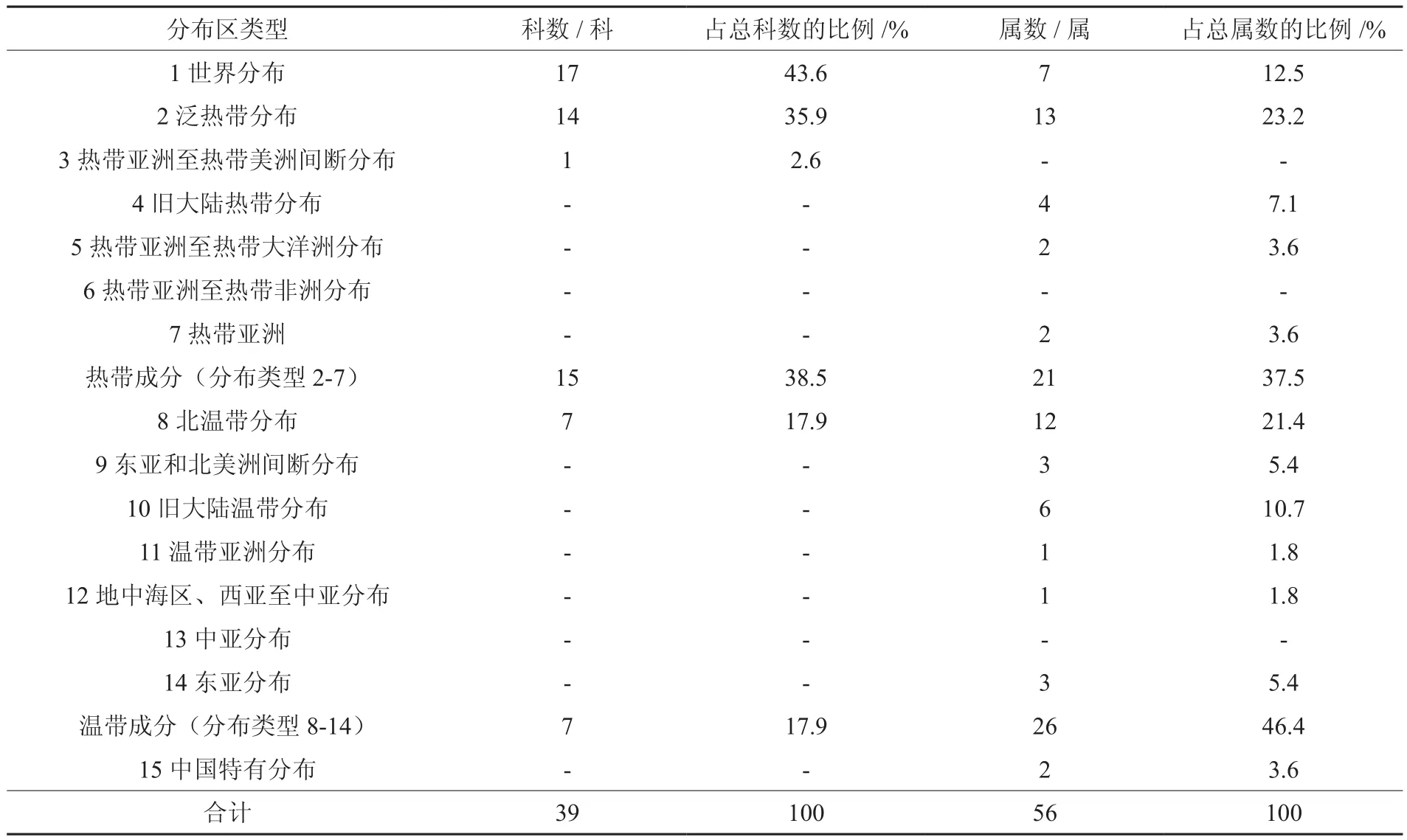
表2 青檀群落种子植物属的分布区类型Table 2 Distribution types of spermatophyte genera in Pteroceltis tatarinowii community
2.3 群落结构
2.3.1 垂直结构
皇藏峪青檀群落的高度在10~18 m之间,成层现象明显,通常可分为乔木层(包括林冠层和亚乔木层)、灌木层和草本层。林冠层高10~18 m,郁闭度为0.6~0.8,除青檀外,常见的树种有黄连木、栓皮栎、五角枫等(见表3)。亚乔木层一般高6~9 m,盖度15%~25%,由青檀、五角枫、栓皮栎、黄连木、小叶朴、鸡桑、黄檀Dalbergia hupeana、栾树Koelreuteria paniculata组成。灌木层高1~4 m,平均盖度15%;该层植物种类较丰富,共出现24种植物,但大都是乔木的幼树,灌木种类仅9种;灌木层的主要构成物种有青檀、扁担木、牡荆、小叶女贞、楝树、圆叶鼠李、五角枫、栓皮栎。草本层高0.6 cm,盖度15%~30%,共出现46种植物,常见的物种有野青茅Deyeuxia arundinacea、求米草Oplismenus undulatifolius、青绿苔草Carex leucochlora、斑叶堇菜Viola variegata、狗牙根Cynodon dactylon、狗尾草Setaria viridis、大油芒Spodiopogon sibiricus以及一些乔灌木的幼苗。此外,群落中共出现7种藤本植物,主要分布于灌草层,常见的种类有太行铁线莲Clematis kirilowii、紫藤Wisteria sinensis、络石Trachelospermum jasminoides。
2.3.2 种群的径级结构
优势种群是群落的重要组成部分,其结构不仅对群落结构有直接影响,而且能客观地体现群落的发展趋势[21]。根据树种的重要值(表3),皇藏峪青檀林乔木层的优势树种有青檀、黄连木、栓皮栎和五角枫4种。按照种群的径级结构,可将上述4种优势树木种群分为两种类型,即增长型种群和稳定型种群(图1)。青檀种群由实生和萌生两类个体组成,从总体上看,种群内的个体在各个径级都有分布,但小径级的个体数占绝对优势,第1、2径级的个体数占总个体数的94.2%,种群的后续资源充足,种群增长趋势明显。但同时,受人为干扰的影响,青檀种群中还存在着大量的萌生个体,这些个体主要集中于第2径级,数量虽多,但生长发育成为较大径级的个体却极少。
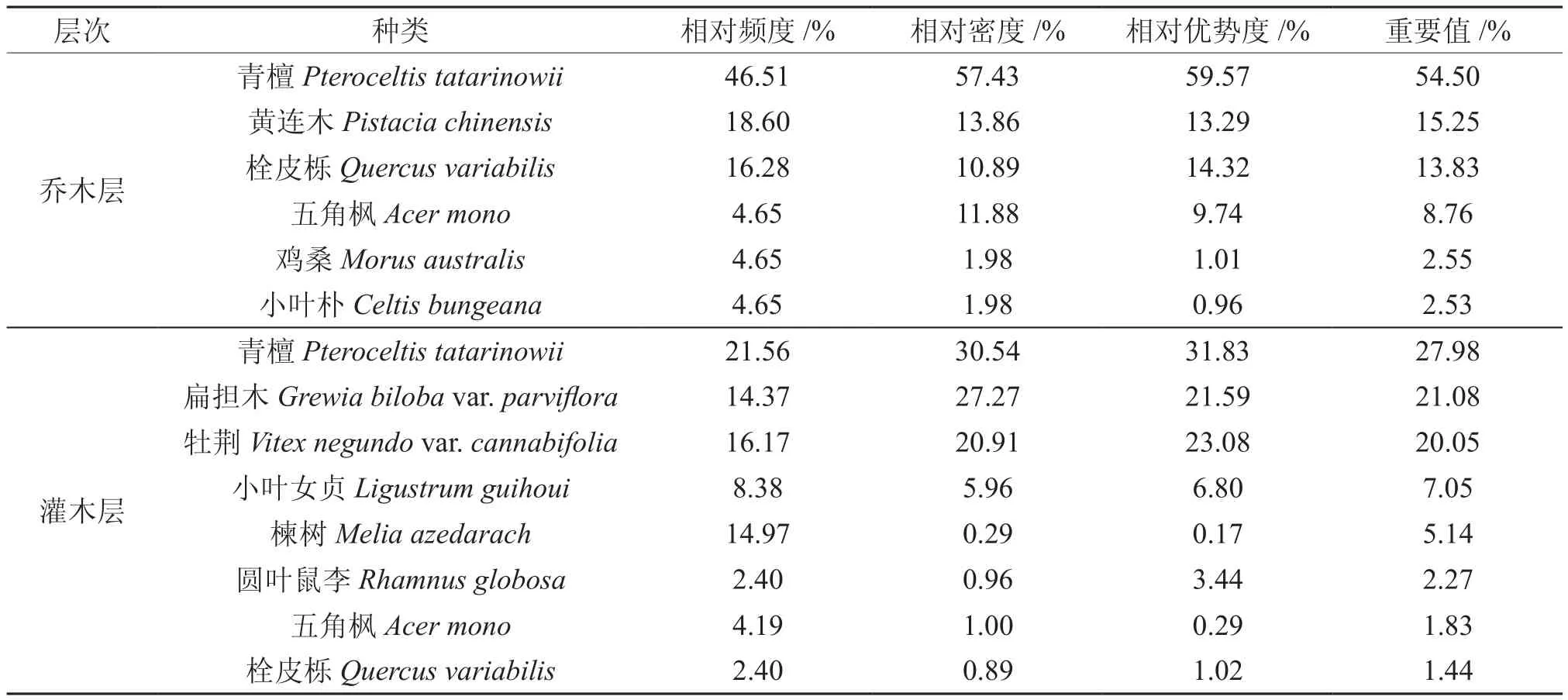
表3 青檀群落主要物种的重要值Table 3 The important values of main species in Pteroceltis tatarinowii community
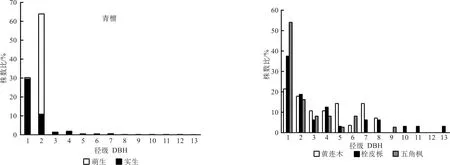
图1 主要乔木种群的径级结构Fig. 1 DBH class distribution of main trees population
青檀群落内黄连木的最大胸径小于28 cm,其种群内的个体分布于前8个径级,各径级的个体数分布较均匀,第1径级的幼苗数最多,但仅占总个体数的21.4%;第6径级树木的个体数最少,占3.6%;最大径级(第8径级)的个体数占7.1%,种群呈稳定的发展趋势。栓皮栎和五角枫2种群的径级结构中都有个别径级个体的缺失,但小径级个体数都占有较高的比例,第1、2径级的个体数分别占个体总数的56.3%(栓皮栎)和70.3%(五角枫),而其它径级的个体数较少,表明种群后续更新的个体充足,种群呈增长的发展趋势。
2.4 群落的外貌
青檀群落林相整齐,林木多数株丛生;落叶成分占绝对优势,常绿半常绿成分共5种(占总种数的8.2%),无常绿乔木,林分季相变化明显。根据Raunkiaer的生活型分类标准,可将皇藏峪青檀林中出现的61种植物的生活型划分为5大类(见表4)。其中,高位芽植物种类最多,有28种,占总种数的45.9%,是青檀群落中占绝对优势的生活型;地面芽植物次之,有22种,占总种数的36.1%;地上芽和地下芽植物各有5种,分别占总种数的8.2%;一年生植物最少,仅1种,占1.6%。可见,青檀林的植物生活型谱以高位芽植物为主,但大高位芽植物缺失,地面芽植物占较大比例,反映了该区生长季水热充足、冬季寒冷的暖温带气候特点。在高位芽植物中,大高位芽植物没有出现;中高位芽植物种类最丰富,有10种,占总种数的16.4%,包括林冠层的青檀、五角枫、栓皮栎、小叶朴等;小高位芽植物和矮高位芽植物各7种,分别占11.5%;藤本高位芽植物4种,占6.5%,反映了我国暖温带落叶阔叶林缺乏高于25m大高位芽植物的特点[22]。

表4 青檀群落的生活型谱†Table 4 The life-form spectrum in Pteroceltis tatarinowii community
4 结论与讨论
皇藏峪自然保护区3块青檀林样地共出现61种植物,分属39科56属,其中单属科和单种属占绝对优势,表明该群落生存环境单一、物种组成复杂多样。群落的组成物种中,热带和温带分布的科、属均占有较大比例,但以温带分布属种居多,表明该群落植物区系成分复杂,温带性质明显,且与热带和温带均有密切的联系,具有由温带区系向亚热带区系过渡的特点[16]。
青檀林的物种组成中落叶成分占绝对优势,常绿成分稀少,无常绿乔木,林分季相变化明显。林内植物以高位芽植物为主、地面芽植物次之,群落的植物生活型谱与秦岭北坡的落叶阔叶林极其相似[23-25],反映了该区温暖湿润、冬季寒冷的气候特点。同时,该区青檀群落的植物生活型谱与广西木论[2]、河南宝天曼[11]两地的青檀群落具有相同的特征,即高位芽植物占绝对优势、地面芽植物占较大比例,反映了青檀群落分布地水热条件的相似性。
青檀群落垂直结构简单,地上成层现象明显,可分为乔木层、灌木层和草本层,青檀在各层中均占明显优势;藤本植物较丰富,主要分布于灌、草层。乔木层的优势树种有青檀、黄连木、栓皮栎和五角枫,这些树种的种群均以小径级个体占优势,除黄连木种群为稳定型种群外,其余3种乔木种群都呈增长的发展趋势,表明该群落具有较强的自我更新和自我维持能力。此外,青檀群落中还存在着大量的青檀萌生个体,它们以第2径级个体为主,多为数株丛生,集中分布于灌木层。
皇藏峪自然保护区的青檀林具有明显的暖温带落叶阔叶林特征,其组成物种复杂多样,垂直结构明显,青檀的优势突出,主要树木种群多呈增长型,群落的自我维持能力较强,稳定性较高。青檀在群落各层中均占有明显的优势,呈现出了较强的生存发展潜力,说明青檀对干旱的石灰岩生境具有良好的适应能力[15],皇藏峪自然保护区的石灰岩山地较适宜于青檀林的分布。但值得注意的是,青檀是具有较强萌生能力的树种,萌生个体对林下个体的生长和群落的持续稳定具有较大的负面影响,而人为的干扰,尤其是保护区的森林管理作业(如砍割灌草、清除枯枝落叶等)是造成青檀个体大量萌生的主要原因,因此,合理调控各项森林管理措施,有效保护林下植被,对维持现有青檀林的稳定具有重要意义。
[1] 杨成华, 安和平, 方小平. 石灰岩山珍稀树种——青檀[J]. 贵州林业科技, 1995, 23(2): 8-11.
[2] 覃文更, 韦国富, 谭卫宁. 广西木论自然保护区青檀群落特征及其多样性研究[J]. 广西林业科学, 2004, 33(3): 126-129.
[3] 傅松玲, 李宏开. 琅琊山青檀天然林群落特征及发展前景探讨[J]. 经济林研究, 1997, 15(1): 13.
[4] 裴海潮. 河南省青檀树种资源分布及采种育苗技术研究[J].河南林业科技, 2007, 27(3): 39-40.
[5] 方升佐, 朱 梅, 唐罗忠, 等. 不同种源青檀种子的营养成分及种子活力的差异[J]. 植物资源与环境, 1998, 7(2): 16-21.
[6] 李光友, 方升佐, 吕家驹, 等. 立地条件对青檀人工林生物生产力及檀皮产量的影响[J]. 南京林业大学学报, 2001, 25(4):49-53.
[7] 洑香香, 方升佐, 汪红卫, 等. 青檀一年生播种苗年生长规律的研究[J]. 南京林业大学学报:自然科学版, 2001, 25(6): 11-14.
[8] 方升佐. 青檀的栽培及檀皮采集加工技术[J]. 林业科技开发,1996, (4): 40-42.
[9] 刘桂华. 青檀耐荫性的初步研究[J]. 经济林研究, 1994, 14(2):7-10.
[10] 许冬芳, 崔同林. 青檀的开发利用[J]. 中国林副特产, 2005,76(3): 64.
[11] 王文静, 何雅蔷. 宝天曼自然保护区青檀林结构特征与物种多样性研究[J]. 河南农业大学学报, 2001, 35(4): 364-367.
[12] 张川红, 郑勇奇, 宗亦臣, 等. 暖温带不同地区青檀形态特征分析[J]. 林业科学研究, 2008, 21(5): 737- 741.
[13] 洑香香, 方升佐, 杜 艳. 青檀种子休眠机理及发芽条件[J].植物资源与环境学报, 2002, 11(1): 9-13.
[14] 江苏省植物研究所编. 江苏植物志(下册)[M]. 江苏:江苏科学技术出版社, 1982:63.
[15] 张兴旺. 珍稀植物青檀的繁殖生物学及其种群结构的研究[D]. 合肥: 安徽师范大学, 2007.
[16] 谢中稳, 蔡永立, 周良骝, 张福林. 安徽皇藏峪自然保护区的植物区系和森林植被[J]. 武汉植物学研究, 1995, 13(4): 310-316.
[17] 郭传友, 任 超. 安徽皇藏峪自然保护区青檀种群的初步研究[J]. 淮北煤炭师范学院学报, 2006, 27(1): 54-57.
[18] 吴征镒, 周浙昆, 李德铢, 等. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 2003, 25(3): 245-257.
[19] 吴征镒. 世界种子植物科的分布区类型系统的修订[J]. 云南植物研究, 2003, 25(5): 535-538.
[20] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究,1991(增刊Ⅳ):1-139.
[21] 达良俊, 杨永川, 宋永昌. 浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J]. 植物生态学报, 2004,28(3): 376-384.
[22] 高贤明, 陈灵芝. 植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析[J]. 植物学报, 1998, 40(6): 553-559.
[23] 李 博 , 杨 持 , 林 鹏 . 生态学 [M]. 北京 : 高等教育出版社 ,2000:127.
[24] 梁文斌,谢碧霞,巫 涛,等.南岳栓皮栎群落特征及多样性分析[J].中南林业科技大学学报,2011,31(9):55-59.
[25] 梁文斌,谢碧霞,巫 涛,等.南岳小红栲的群落学特征研究[J].中南林业科技大学学报,2011,31(10):15-20.
Community characteristics of Pteroceltis tatarinowii forests in Huangcangyu natural reserve of Anhui province
YOU Hai-mei1, YU Fa-zhan1, YAN Сhuan-hai2
(1. School of Urban and Environment, Jiangsu Normal University, Xuzhou 221116, Jiangsu, Сhina;2. School of Environmental and Municipal Engineering, Qingdao Technological University, Qingdao 266033, Shandong, Сhina)
To understand the developing state and to effectively conserve the community of naturalPteroceltis tatarinowiiforests in Huangcangyu natural reserve, the characteristics of the species composition, community structure and physiognomy were studied based on sampling survey method. The results show that there were 61 vascular plants in the community belonging to 56 genera and 39 families. The analysis of fl oristic geographical elements indicate that both tropical and temperate elements occupied high percentage respectively, however, the contribution of genera and species belonging to temperate areal-types was larger, and the composition of the fl ora appeared transitional character of temperate fl ora to tropical fl ora. The analysis of life form spectrum and leaf characters shows that the physiognomy of the community was characterized by deciduous phanerophytes and the hemicryptophytes had a large contribution.The community vertical structure was simple, which composed of tree layer, shrub layer and herb layer, andPteroceltis tatarinowiiwas dominated in each layer; the forest natural regeneration was good, most populations of main tree species were increasing type, and the community had higher stability. However, many coppice shoots ofPteroceltis tatarinowiicaused mainly by human disturbance (e.g.cutting the shrubs and herbs), were mainly distributed in shrub layer and herb layer with bundle consist of coppice sprouts, affecting the growth of understory plants and the stability of the community. Thus, the vegetation in understory should be conserved effectively.
Pteroceltis tatarinowii; community; species composition; structure; physiognomy; Huangcangyu natural reserve
S718.54
A
1673-923X (2012)05-0086-06
2012-02-04
日本文部省科研基金项目 (16255003);徐州师范大学科研基金项目(10XLA10)
尤海梅(1965-),女,黑龙江哈尔滨人,讲师,博士,主要从事植被生态学、种群生态学研究
[ 本文编校:吴 毅]
