A behavioral ecology approach to traffic accidents: Interspecific variation in causes of traffic casualties among birds
2011-12-25AndersPapellerHelgaErritzJohannesErritz
Anders Pape Møller, Helga Erritzøe , Johannes Erritzøe
(1. Department of Ecology, Systematics and Evolution, CNRS UMR 8079, University of Paris-South, Bâtiment 362, F-91405 Orsay Cedex, France; 2. Taps Old Rectory, DK-6070 Christiansfeld, Denmark)
A behavioral ecology approach to traffic accidents: Interspecific variation in causes of traffic casualties among birds
Anders Pape Møller1,*, Helga Erritzøe2, Johannes Erritzøe2
(1.Department of Ecology, Systematics and Evolution, CNRS UMR8079,University of Paris-South, Bâtiment362,F-91405Orsay Cedex,France; 2.Taps Old Rectory, DK-6070Christiansfeld,Denmark)
Birds and other animals are frequently killed by cars, causing the death of many million individuals per year. Why some species are killed more often than others has never been investigated. In this work hypothesized that risk taking behavior may affect the probability of certain kinds of individuals being killed disproportionately often. Furthermore, behavior of individuals on roads, abundance, habitat preferences, breeding sociality, and health status may all potentially affect the risk of being killed on roads. We used information on the abundance of road kills and the abundance in the surrounding environment of 50 species of birds obtained during regular censuses in 2001-2006 in a rural site in Denmark to test these predictions. The frequency of road kills increased linearly with abundance, while the proportion of individuals sitting on the road or flying low across the road only explained little additional variation in frequency of road casualties. After having accounted for abundance, we found that species with a short flight distance and hence taking greater risks when approached by a potential cause of danger were killed disproportionately often. In addition, solitary species, species with a high prevalence ofPlasmodiuminfection, and species with a large bursa of Fabricius for their body size had a high susceptibility to being killed by cars. These findings suggest that a range of different factors indicative of risk-taking behavior, visual acuity and health status cause certain bird species to be susceptible to casualties due to cars.
Birds; Blood parasites; Flight behavior; Flight distance; Habitat selection; Sociality
The impact of humans on animals has intensified dramatically during the last decades due to increasing human populations and the resulting exploitation of natural resources across the globe (Marzluff et al, 2001).Urbanization has a number of consequences for wild animals because of the longer growing season associated with altered micro-climatic conditions, a change in abundance of habitats, an increase in food abundance and hence population density of specific animals, and proximity of humans that may be problematic for species with low thresholds of fear (Batten, 1973; Klausnitzer, 1989; Gliwicz et al, 1994; Stephan, 1999; Shochat et al, 2006; Møller, 2008a). A major impact of urbanization is the construction of roads that disrupt animal movement, but also affect everyday life of animals due to the presence of cars and their effects. For example, recent research has shown strong effects of noise from traffic on song learning and change in composition and carrier frequency of songs (e. g. Reindt, 2003; Warren et al, 2006). Second, roads can have dramatic effects on the distribution of animals (Reijnen & Foppen, 1991; Underhill & Angold, 2000; Forman & Alexander, 1998). A third consequence is the large number of traffic kills due to cars (Blümel & Blümel, 1980; Bruun-Schmidt, 1994; Dunthorn & Errington, 1964; Erritzøe et al, 2003; Finnis, 1960; Göransson et al, 1978; Haas, 1964; Heinrich, 1978; Hodson, 1960; Hodson & Snow, 1965; Nankinov & Todorov, 1983; Smettan, 1988; Straube, 1988; Vignes, 1984; Wäscher et al, 1988). For example, the total number of birds killed by traffic in Denmark alone (a small country of only 43,000 km2) was estimated to be 3.50 million in the 1960’s and 3.27 million in 1980 (Hansen, 1969, 1982). Therefore, the global estimates must at least be hundreds of millions per year.
While the effect of cars on avian mortality is well described, there is to the best of our knowledge no information about the causes of interspecific variation in such mortality. Superficially, one might assume that all species are equally susceptible to traffic mortality because the recent invention of cars has not allowed adaptation to this cause of mortality. However, there might be behavioral or physiological traits that predispose particular species to elevated risk of mortality. Are road-kills just reflecting a random sample of all species according to their abundance and the probability that individuals sit on the road or fly close to the ground, thereby exposing them to traffic accidents, or do specific characteristics of certain species render them more susceptible to being killed by cars? The objectives of this study were to address to which extent the abundance of road-kills of different species could be explained by ecological factors. To this end we estimated both the frequency of road-kills of different species, but also their expected frequency based on standardized birds censuses.
The aims of this study were to assess risk factors associated with road-kills of birds. As a null hypothesis we assessed the importance of abundance as a predictor of the frequency of road-kills of different species, because abundant species by necessity are more likely to end up being killed by cars. Next, we calculated an index of susceptibility to mortality due to cars by quantifying residual variation for each species. First, we hypothesized that species living in open habitats would have a better ability to escape an approaching car because of a better view of the approaching vehicle. Second, Blumstein (2006) has in a study of the evolutionary ecology of fear suggested that flight initiation distance when approached by humans provides a reliable metric of wariness. Flight distance to an approaching human can be considered a metric of the risk that individuals of a given species are willing to take when encountering a potential predator (Blumstein, 2006). Importantly, Møller et al (2008) found that bird species with relatively short flight distances suffered from higher susceptibility to predation by sparrowhawksAccipiter nisus, and susceptibility of birds to cat predation is also related to flight distance (Møller et al, 2010). Therefore, species with short flight distances should be particularly susceptible to traffic. Species feeding on fast-moving prey may be better at motion detection (Fleishman et al, 1995) and have more acute visual systems (Garamszegi et al, 2002), making it likely that species eating moving prey would be more flighty than herbivores. Hence, species eating moving prey should be less susceptible to traffic than herbivores. Sociality may involve allocation of more time to monitor conspecifics (Roberts, 1988), making social species more vigilant to an approaching human being (Blumstein, 2006), but incidentally also to approaching cars. Third, we predicted that a relatively short duration of the nestling period for a given body size would imply that fledglings had not fully developed when leaving the nest (Starck & Ricklefs, 1998), resulting in an increased risk of mortality due to cars in such species. Fourth, we predicted that malaria and other blood parasites would affect metabolic efficiency of hosts (Atkinson & Van Riper, 1991; Chen et al, 2001), thereby reducing working ability and hence escape ability when a car is rapidly approaching. Thus, we predicted that bird species with a high prevalence of blood parasites, especially intracellular blood parasites that cause erythrocytes to burst and hence give rise to anemia, would be positively related to susceptibility to car fatalities. Finally, we expected that aspects of immunity in young birds would predict susceptibility to car accidents because such measures of immunity would have evolved in response to parasite-mediated selection, causing species with virulent parasites to have particularly large immune defense organs for their body size. Thus, we predicted a positive relationship between susceptibility to car accidents and relative size of the bursa of Fabricius, which is an important immune defense organ involved in production of antibodies and responsible for differentiation of the repertoire of B-cells in young birds (Glick, 1983, 1994; Toivanen & Toivanen, 1987). Many studies have shown that relative size of the bursa of Fabricius is related to parasite impact on avian hosts (e.g., Garamszegi et al, 2003, 2007; Garamszegi & Møller, 2007; Møller, 2009; Møller & Erritzøe, 1996).
In the present study we were able to assess traffic casualties in relation to null expectations based on abundance and flight behavior. In addition, we performed post mortem autopsies of almost all individuals, verifying that they indeed had died from impact with a car. We tested our predictions concerning risk factors associated with traffic fatalities of birds by relying on an extensive data set of road-kills collected by JE and HE, who also conducted standardized bird censuses in the same sites to quantify the abundance of different species. Therefore, the present study assessed risk factors of traffic mortality in birds, because we could compare observed and expected abundance of road-kills of different species and relate such an index of susceptibility to flight behavior, and ecological and life history characteristics. Thus, we were able to assess the relative importance of a range of different predictors of susceptibility to traffic accidents.
1 Materials and Methods
1.1 Road kills
JE and HE biked or walked 10,500 km on a fixed route during 2001-2006 around Taps (55o27’ N, 9o26’ E), Denmark, with an even distribution of these sampling events throughout the year. The habitat of this open farmland area is open fields, with scattered hedges, plantations and houses with gardens. Traffic is not intense because less than 70 cars were recorded on most censuses. Each trip covered at least 6 km, and they found a total of 461 road-killed birds by carefully searching the road and the verges during these trips. All individuals were aged according to standard criteria reported by Svensson (2006). In addition, all individuals were inspected post mortem, and traffic casualty rather than other causes of mortality was ascertained from severe fractures, blood-filled lungs and other signs of a forceful impact between a bird and a hard obstacle (a car). Thus, we could exclude the possibility that the birds recorded had died for other reasons such as collision with overhead wires, poisoning, predation or other causes of death. The species and their abundance are listed in the Appendix.
JE also recorded all individuals seen during the regular trips, carefully recording whether an individual was sitting on the road or road verge, and whether an individual flew at a height of less or more than 2 m, as judged from our own height. We used this information to estimate the proportion of individuals of each species that was sitting on the road and the proportion of individuals that flew below a height of 2 m as indicators of elevated risk of impact with cars.
A total of 25 of 249 sparrowsPassersp. (10%) could not be assigned to species with certainty, and we attributed these unidentified sparrows to house sparrowsPasser domesticusand tree sparrowsPasser montanusaccording to the relative abundance of these two species as road kills of actually identified individuals. However, the results remained unchanged if these 25 unidentified individuals were excluded from the analyses.
1.2 Bird censuses
JE conducted standardized point counts (Bibby et al, 2005) to quantify the abundance of birds during the breeding season and the winter along the roads where road-kills were recorded. In brief, the point counts consisted of 5 minutes being spent per point looking and listening for birds at any distance from the point, while recording these, with 100 m between individual census points, using a total of 20 points per census. Total abundance during the breeding season was estimated as the sum of the seven annual point count censuses made between 2001 and 2006 during the breeding season between 25 May and 1 June. Two winter censuses were conducted on 5 January 2005 and 5 January 2006. We obtained an estimate of total abundance of each species by adding the number of breeding season observations to the number of winter observations multiplied by 3.5, to account for the fact that there were 3.5 times as many breeding season censuses as winter censuses. The bird count results were repeatable among censuses during the breeding season (one-way ANOVA based on the log10-transformed counts:F= 3.72,df= 33,170,P< 0.0001) and winter (F= 4.66,df= 7,8,P= 0.023), implying that the abundance of different species was consistent among years. The results of these censuses are reported in the Appendix.
1.3 Ecological variables
We scored the main breeding habitats as open grassland (a score of 0), bushes (a score of 1), or trees (a score of 2), relying on information in Cramp & Perrins (1977-1994).
During March-August 2006-2007 APM estimated flight distances for birds in Ile-de-France and Bretagne, France and Northern Jutland, Denmark, using a standard technique developed by Blumstein (2006). Estimates of flight distances are highly consistent among countries, as reported by Møller (2008a, b), hence justifying the use of flight distance data from another population than that used for studying traffic mortality. In brief, when an individual bird had been located while resting, foraging, preening or singing, APM moved at a normal walking speed towards the individual, while recording the number of steps (which approximately equals the number of meters). The distance at which the individual took flight was defined as the flight distance, while in accordance with Blumstein (2006) we defined starting distance as the distance from where the observer started walking until the position of the bird. If the individual was positioned in the vegetation, the height above ground, when APM started walking towards it, was recorded to the nearest meter. While recording these distances, APM also recorded date and time of day, age and the sex of the individual if external characteristics allowed sexing with binoculars. Juveniles were only included when they were fully independent and not fed by adults. Flight distance was estimated as the squareroot of the sum of the squared horizontal distance and the squared height (Blumstein, 2006).
All recordings were made during the breeding season, when most individuals are sedentary, thus preventing the same individual from being recorded in different sites, with each site being more than 100 m apart (equaling a territory of 3.1 ha), and for larger species such as herons and corvids APM used a distance of 500 m (equaling a territory of 78.5 ha) because of their larger territories. Using only breeding birds may cause another problem if territorial individuals are less likely to have long flight distances because they are reluctant to cross territory boundaries. However, we can dismiss this alternative explanation because an analysis of 24 species (based on 2018 observations), assessed during winter (December 2006 - February 2007) and summer revealed a strong positive relationship between the two sets of estimates (weighted regression:F= 41.50,df= 1, 3,r2= 0.64,P< 0.0001).
We assessed the spatial and temporal consistency in estimates of mean flight distances in three ways and found high degrees of consistency (Møller, 2008a,b).
Breeding sociality was quantified as colonial (a score of 1) when individuals used territories that only contained a nest site, and otherwise solitary (a score of 0), using information in Cramp & Perrins (1977-1994).
We extracted information on duration of the incubation and the nestling period from Cramp & Perrins (1977-1994), relying on the value based on the largest sample size if more than a single value was reported. If a range of values was reported, we used the mean of the extreme values. Likewise, we extracted mean body mass from the breeding season from Cramp & Perrins (1977-1994), using the largest sample if more data were reported, and using the mean of means for adult males and adult females, if masses were reported separately for the two sexes.
We used information on prevalence of four genera of blood parasites (Leucocytozoon,Haemoproteus,Plasmodium,Trypanosoma), assuming that this would reflect prevalence in the population, as done in previous studies (e. g. Møller & Nielsen, 2007). We also extracted information on the number of individuals examined for each of the host species. In total, the study was based on examination of infection level of 16,995 individual juvenile and adult hosts belonging to 263 species based on blood smears, with a range from 2 to 1539 individuals each for the species [see Møller & Nielsen (2007) for a complete list of sources]. Most of the blood parasite information derived from Northern Europe, where the study of road-kills was also conducted, providing a high degree of overlap between the geographical location of study sites for parasites and car casualties. Scheuerlein & Ricklefs (2004) have shown that prevalence estimates are repeatable across study sites, despite considerable variation among sites and sampling periods. Such variation among sites and sampling periods would make any investigation of relationships between prevalence and ecological variables conservative.
We used information on the mass of the bursa of Fabricius from Møller et al (2005) and Garamszegi et al (2007).
The entire data set is reported in the Appendix.
1.4 Statistical methods
We estimated a susceptibility index to quantify likelihood for individuals of a given species to be killed by traffic, as log10-transformed number of road-kills minus log10-transformed (number of individuals observed of that species divided by the total number of individuals of all species) multiplied by the total number of road-kills). To avoid problems with species having an abundance of road-kills or an overall abundance of zero, we added one to all observations.
Nestling period, mass of bursa of Fabricius, body mass and flight distance were log10-transformed, while the proportion of individuals sitting on the road, the proportion of individuals that flew at a height of less than 2 m and prevalence of blood parasites were square-root arcsine-transformed before analysis.
Most statistical approaches assume that each data point provides equally precise information about the deterministic part of total process variation, i.e. the standard deviation of the error term is constant over all values of the predictor variables (Sokal & Rohlf, 1995). We weighted each observation by sample size (the number of individuals killed by traffic) in order to use all data in an unbiased fashion, thereby giving each datum a weight that reflects its degree of precision due to sampling effort (Draper & Smith, 1981; Neter et al, 1996). Comparative analyses may be confounded by sample size if sampling effort is important, and if sample size varies considerably among taxa. In order to weight regressions by sample size in the analysis of contrasts, we calculated weights for each contrast by calculating the mean sample size for the taxa immediately subtended by that node in the phylogeny (Møller & Nielsen, 2007).
We calculated an estimate of effect size as the Pearson product-moment correlation, using the equations in Rosenthal (1991, pp. 73-74), relying on Cohen’s (1988) conventions wherer= 0.1 equals a small effect,r= 0.3 an intermediate effect andr= 0.5 equals a large effect).
The three most common species accounted for 75% of all road-kills, implying that these species may have a disproportionate influence on the conclusions. We re-did all analyses after exclusion of these three common species, but all conclusions remained unchanged, suggesting that the three most common species did not bias the findings.
1.5 Comparative analyses
We controlled for similarity in frequency of traffic casualties among species due to common ancestry by calculating standardized independent linear contrasts (Felsenstein, 1985), using the computer program CAIC (Purvis & Rambaut, 1995). Standardization of contrast values was checked by examination of absolute values of standardized contrasts versus their standard deviations (Garland et al, 1992). Plotting the resulting contrasts against the variances of the corresponding nodes revealed that these transformations made the variables suitable for regression analyses. In cases where extreme residuals were recorded, we tested for the robustness of the conclusions by excluding contrasts with studentized residuals greater than 3.00 (Jones & Purvis, 1997). Likewise, we ranked the independent variable to test if the conclusions remained unchanged (Møller & Birkhead, 1994), and in no case did this procedure give rise to conclusions different from those obtained with the contrast values. Ranking provides a very conservative test of a hypothesis, and robustness of findings to ranking of the independent variable thus suggests that distributions of variables are not a confounding factor leading to specific conclusions.
We constructed a composite phylogeny of the 50 species investigated to calculate standardized linear contrasts, mainly based on Sibley & Ahlquist (1990), combined with information from more recent sources (Blondel et al, 1996; Barker et al, 2001, 2004; Crochet et al, 2000; Helbig & Seibold, 1999)(Fig. 1). Because this composite phylogeny was derived from different studies using different phylogenetic methods and different molecular markers, consistent estimates of branch lengths were unavailable. Therefore, we considered branch lengths to be equal in the analyses (this is equivalent to an assumption of a punctuated model of evolution (Purvis & Rambaut, 1995)). We used MacClade to construct the phylogeny and make inferences about the ancestral state of susceptibility to traffic (Maddison & Maddison, 1992).
Regressions of standardized linear contrasts were forced through the origin because the comparative analyses assume that there has been no evolutionary change in a character when the predictor variable has not changed (Purvis & Rambaut, 1995). We used multiple regression to find the best-fit model, using the software JMP (Anon, 2000). The best-fit model was determined using Akaike’s information criterion as an estimate of the improvement in fit for addition of variables (Burnham & Anderson, 2002). There was no evidence of collinearity between variables (Tabacknik & Fidell, 1996).
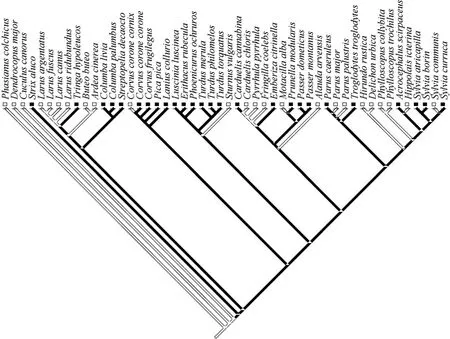
Fig. 1 Phylogeny of species of birds analyzed for road-kills
Information was missing for some species for certain variables, causing sample sizes to differ slightly among analyses.
Values reported are means (SE).
2 Results
The number of individuals killed varied among 50 bird species from 0 to 199, with a mean = 9.2, median = 2.0,SE= 4.4. The three most abundant species were house sparrow (199 individuals), blackbirdTurdus merula(95) and tree sparrow (49). The total abundance during censuses of the 50 species varied from 0 to 261 individuals, with a mean = 12.5, median = 2.5,SE= 5.4. The frequency of traffic kills increased significantly with abundance in a linear fashion, explaining 42% of the variance (Fig. 2), and the increase was significant both for analyses based on species-specific data and independent contrasts (Tab. 1). The susceptibility index varied from -1.235 to +0.699 with a mean of 0.005 (SE= 0.061), not differing significantly from zero (onesample t-test,t= 0.08,df= 49,P= 0.94). The distribution of the relative frequency of road-kills on the phylogeny showed that the ancestral state among the species studied was a low frequency of traffic casualties (Fig. 1). There were many transitions between different states (Fig. 1), suggesting that ecological factors associated with a relatively high frequency of road-kills were labile.
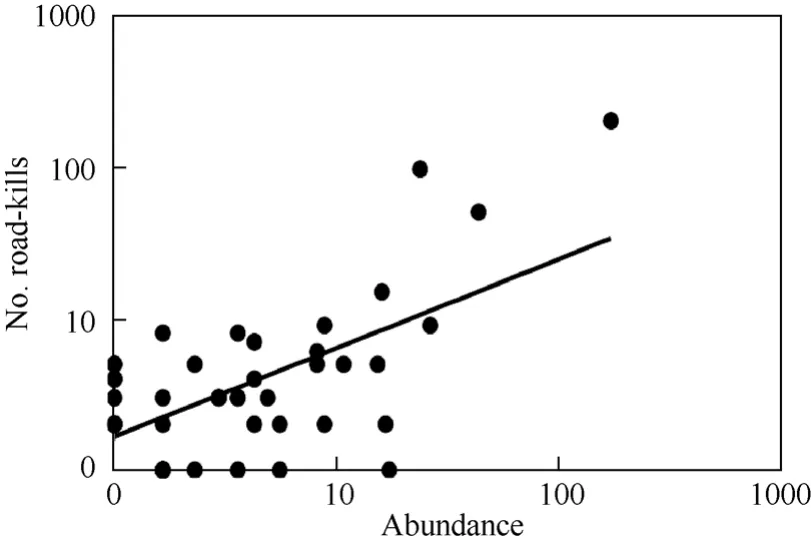
Fig. 2 Frequency of traffic kills in different species in relation to their abundance
The proportion of individuals flying below 2 m ranged from 0 to 1 among species, with a mean value of 0.36 (SE= 0.05). The proportion of individuals on the road varied from 0 to 1 among species, with a mean value of 0.31 (SE= 0.05). These two variables were not significantly correlated (Pearsonr= 0.06,t= 0.44,df= 48,P= 0.66). The proportion of individuals on the road explained a significant amount of variation in road-kills (F= 6.65,df= 1,48,P= 0.013, slope (SE) = 0.32 (0.12), effect size = 0.35), while that was not the case for the proportion of individuals flying below 2 m (F= 0.38,df= 1,48,P= 0.54, slope (SE) = 0.08 (0.12), effect size = 0.09). Neither variable explained a significant amount of variation in susceptibility to traffic (proportion of individuals on road: partialF= 0.81,df= 1,47,P= 0.37, effect size = 0.13; proportion of individuals flying lower than 2 m: partialF= 0.07,df= 1,47,P= 0.80, effect size = 0.04). Flight distance did not significantly predict susceptibility to traffic in a model that included body mass (partialF= 1.10,df= 1,42,P= 0.30, effect size = 0.16).

Tab. 1 Linear regression of log-transformed number of road-kills for different species of birds in relation to log-transformed abundance
Age did not affect susceptibility to traffic because the proportion of yearlings was not a significant predictor in a model that included body mass as an additional predictor variable (partialF= 0.14,df= 1,33,P= 0.71, effect size = 0.06).
Susceptibility to traffic increased from open habitats and bushes to trees in analyses of species-specific data, but not in analyses of contrasts (Tab. 2 models A-B).
Breeding sociality reduced susceptibility to traffic, after inclusion of body mass as an additional predictor variable (Fig. 3A; partialF= 11.52,df= 1,46,P= 0.0014, effect size = 0.45). In contrast, food did not significantly predict susceptibility to traffic, after inclusion of body mass as an additional predictor variable (partialF= 0.00,df= 1,46,P= 0.99, effect size = 0.01).
Bird species with a long nestling period for their body size were less susceptible to traffic, and this effect was independent of the effect of habitat in analyses of species-specific data and contrasts (Tab. 2 Models A-B). In contrast, the duration of the incubation period was not a significant predictor (results not shown). In addition, susceptibility increased with body mass, although only in analyses of species-specific data (Tab. 2 Models A-B), implying that large species were more susceptible than small species in species-specific analyses.
The prevalence of two genera of blood parasites,LeucocytozoonandPlasmodium, were both positively related to susceptibility to traffic, with similar effects in analyses of species-specific data and independent contrasts (Tab. 2 Models C-D; Fig. 3B). In contrast, the prevalence ofHaemoproteusandTrypanosomawas not a significant predictor (results not shown for simplicity).
The size of the bursa of Fabricius was positively related to susceptibility to traffic, both in analyses of species-specific data and independent contrasts (Tab. 2 Models E-F; Fig. 3C). This effect was independent of body mass.
Finally, we developed best-fit models that included all the predictor variables. An analysis of speciesspecific data revealed a model that explained 60% of the variance (Tab. 3 Model A). There were statistically significant effects of flight distance after inclusion of the other variables of interest (Fig. 3D; negative relationship between distance and susceptibility), breeding sociality (solitary species having greater susceptibility than colonial species), and mass of bursa of Fabricius (species with larger bursa being more susceptible) (Tab. 3 Model A). A model based on independent contrasts revealed intermediate to strong effects of flight distance (negative relationship between distance and susceptibility), breeding sociality (solitary species having greater susceptibility than colonial species), food (species eating live prey being less susceptible than herbivores), prevalence of
Plasmodium(species with higher prevalence being more susceptible), bursa of Fabricius (species with larger bursa being more susceptible), and body mass (larger species being less susceptible) (Tab. 3 Model B).
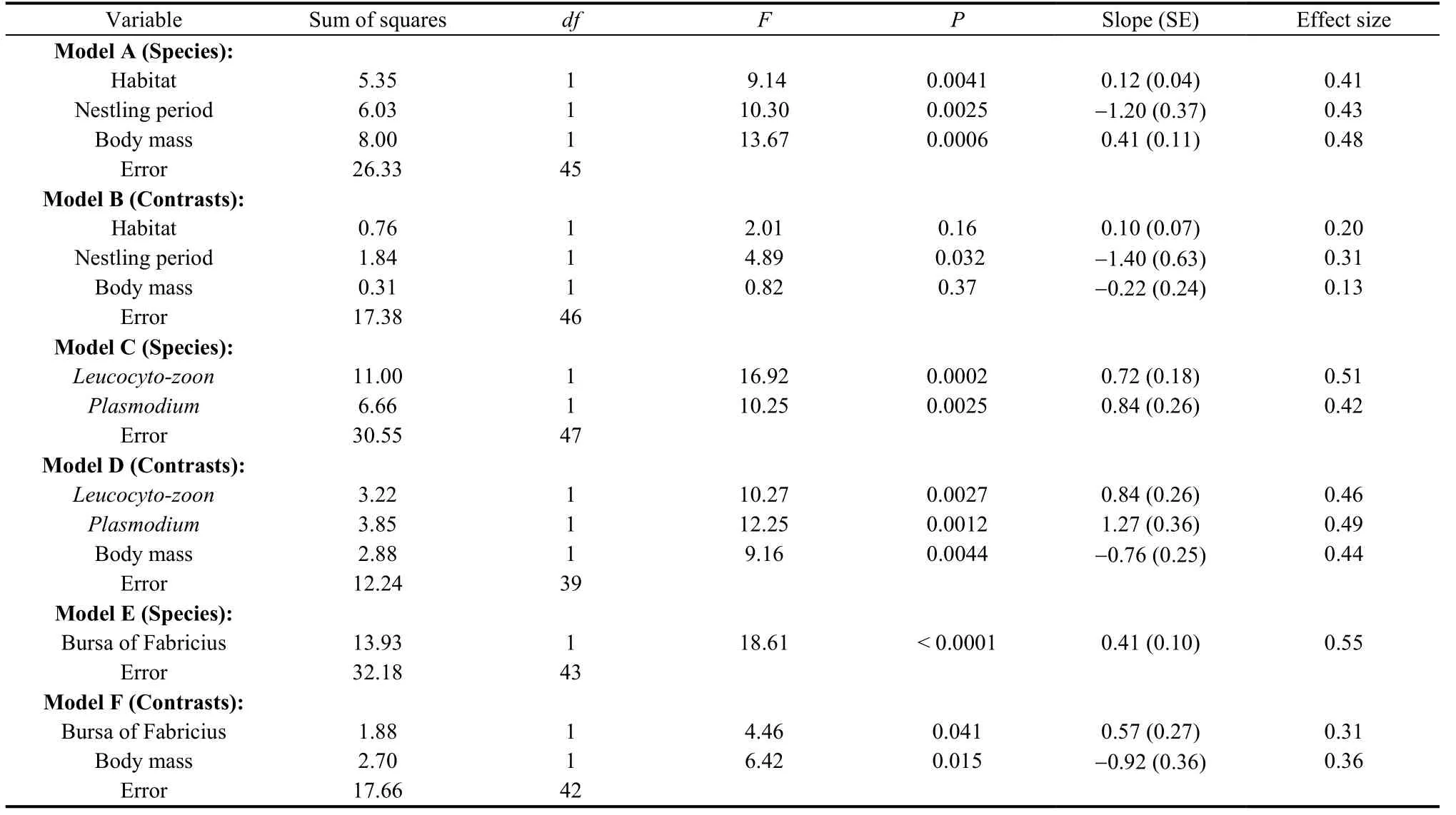
Tab. 2 Best-fit models of the relationship between susceptibility to mortality due to traffic and (models A-B) habitat, logtransformed nestling period and log-transformed body mass, (models C-D) square-root arcsine-transformed prevalence of Leucocytozoon and Plasmodium and log-transformed body mass, (models E-F) log-transformed mass of bursa of Fabricius and log-transformed body mass
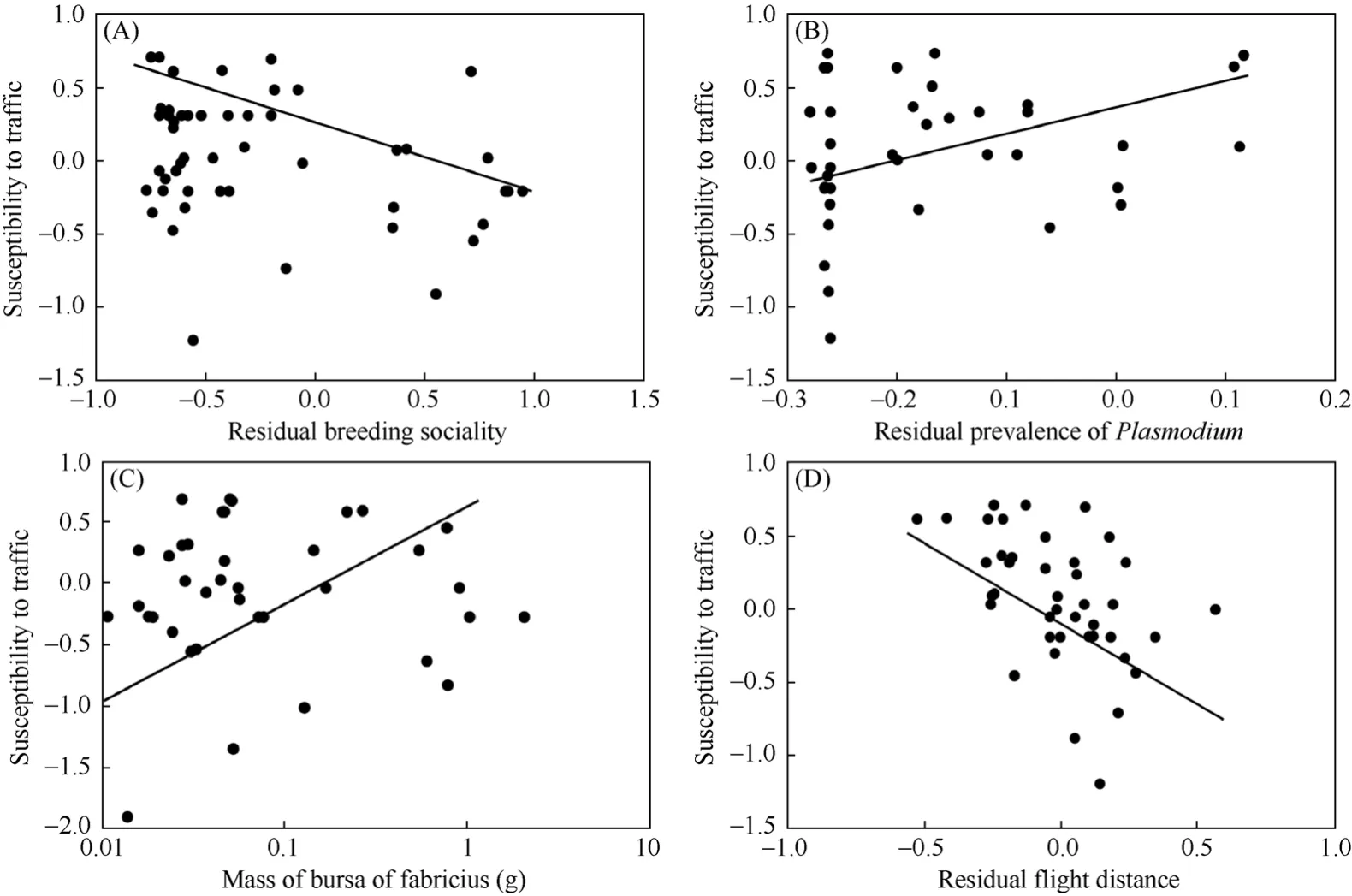
Fig. 3 Susceptibility to traffic death in different species of birds in relation to (A) breeding sociality, (B) relative prevalence of Plasmodium (after adjusting for prevalence of Leucocytozoon and body mass), (C) mass of the Bursa of Fabricius (g), and (D) flight distance (after adjusting for breeding sociality and mass of bursa of Fabricius)
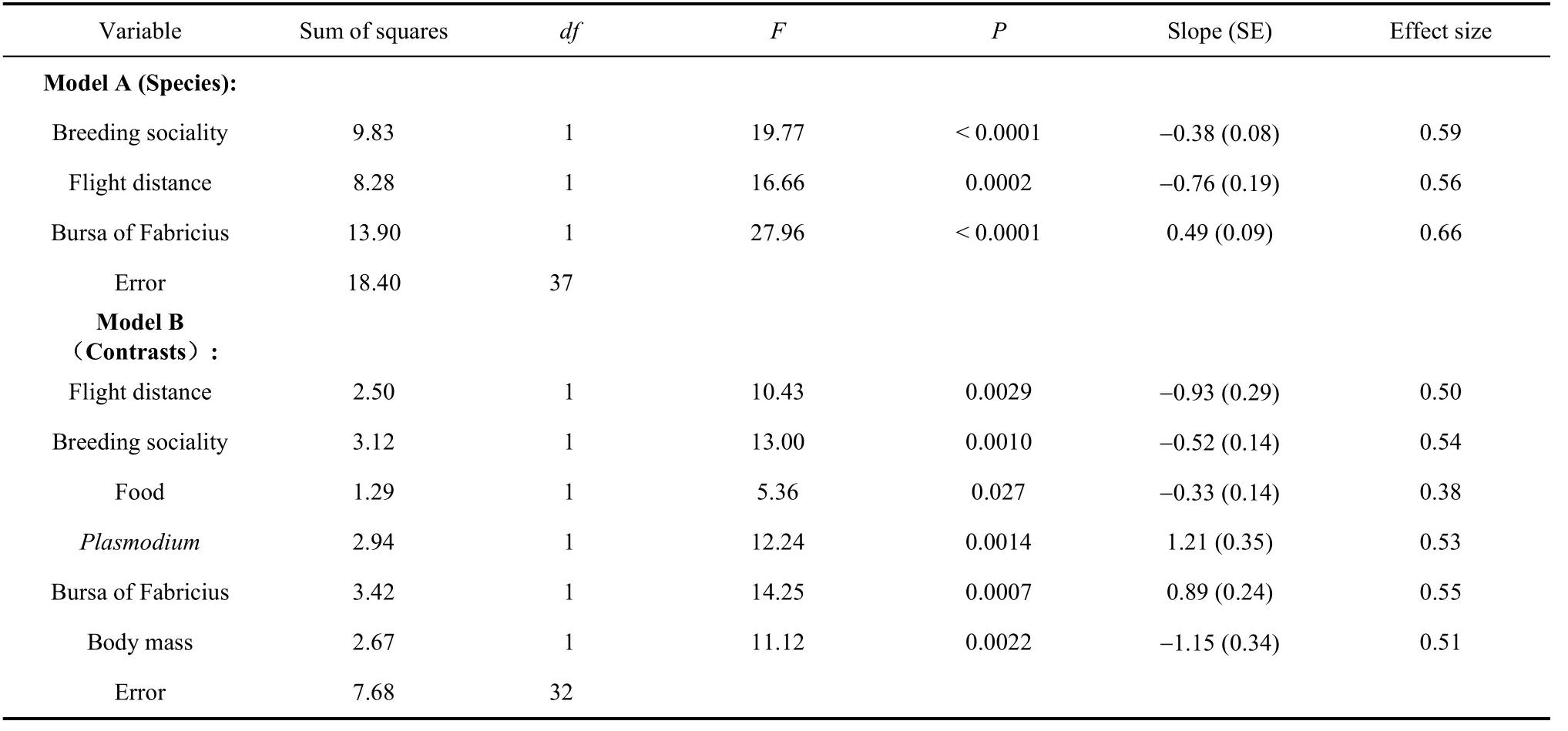
Tab. 3 Best-fit models of the relationship between susceptibility to mortality due to traffic and habitat, log-transformed nestling period, square-root arcsine-transformed prevalence of Leucocytozoon and Plasmodium, log-transformed mass of bursa of Fabricius and log-transformed body mass
3 Discussion
We provide evidence of avian mortality due to traffic directly reflecting the abundance of different species, but also phenotypic characteristics relating to flight behavior, visual acuity and health status. The null hypothesis investigated was that the abundance of traffic casualties simply reflected the abundance of different species, but also their behavior, with species with many individuals sitting on the road or flying close to the ground experiencing a high risk of mortality due to traffic. Abundance and to a small extent the proportion of individuals that were sitting on the road explained variation in traffic casualties. In addition, risk-taking behavior when approached by a human being estimated in terms of flight distance, breeding sociality (solitary species run higher risks), prevalence of the blood parasites of the genusPlasmodium(species with high prevalence run higher risks), and the size of the bursa of Fabricius (species with large immune defense organs run higher risks) all predicted susceptibility to traffic. We will briefly discuss these findings.
Mortality among birds due to traffic may constitute a non-negligible fraction of overall mortality, and Svensson (1998) suggested that this factor is seriously under-estimated. Hansen (1982) estimated that 3.27 million birds died on roads in Denmark in 1980-1981. Given that the total population of birds, including fledglings, consists of 16.29 million pairs (based on the mean estimates reported by Grell (1998)), and assuming that mean clutch size is 4.5 eggs (the mean for 512 species breeding in the Western Palearctic (Cramp & Perrins, 1977-1994), the mean number of clutches per year being 1.6 (the mean for 512 species breeding in the Western Palearctic (Cramp & Perrins, 1977-1994)), and mean nest predation rate being 24% for 72 species (Cramp & Perrins, 1977-1994)), then the total population can be estimated as (16.29 million pairs × 2 individuals per pair) + (16.29 million pairs ×4.5 eggs per clutch × 1.6 clutches per year × 0.76) = 121.72 million individuals. Therefore, mortality due to traffic can be estimated as (3.27 million / 121.72 million), or 2.7%. Given that mean adult mortality rate for 172 bird species from the Western Palearctic (Cramp & Perrins, 1977-1994) was 38%, mortality due to traffic accounted for 2.7% / 38% = 7.1% of overall mortality. This proportion is bound to be an over-estimate because juvenile mortality is likely to be greater than 38%. However, the conclusion remains that avian mortality due to traffic constitutes a non-negligible fraction of overall mortality in the order of 5%-10% of overall mortality. Whether this has any effect on population size will depend on whether mortality caused by traffic acts in a density-dependent manner. To the best of our knowledge this provides the first estimate of the relative importance of traffic casualties for overall mortality in any group of animals.
The null hypothesis for traffic casualties is that risk of mortality should reflect the abundance of different species and their flight behavior. Abundance explained 42% of the variance in traffic mortality among species. Surprisingly, we found little evidence of flight behavior affecting the abundance of traffic casualties. There was a weak positive relationship between the proportion of individuals of different species being recorded on the road and road-kills, while the proportion of individuals flying at a height of less than 2 m did not predict traffic casualties. This leaves 60% of the variance to be explained by other factors such as sampling effort, ecological factors and other sources of variation.
Blumstein (2006) suggested in his hypothesis about the evolutionary ecology of fear that flight distance should provide a reliable metric reflecting the risk that individuals are willing to take when trading foraging against flight. Studies of flight distance in birds have indicated that urban birds take greater risks than rural conspecifics, as reflected by shorter flight distances (Møller, 2008a). A number of factors hypothesized to provide increased visual acuity such as sociality and reliance on live prey may also allow individuals to escape when approached by a car. We found clear evidence suggesting that flight distance and breeding sociality both predicted susceptibility to traffic, and that was also the case in analyses of contrasts.
Risk of mortality due to traffic was elevated in bird species with high prevalence of blood parasites of the genusPlasmodium. While we have no information whether individuals killed by cars were infected by blood parasites, we can conclude that species that had high prevalence ofPlasmodiumsuffered disproportionately from traffic mortality. We have no a priori explanation why there were no effects for the blood parasite generaHaemoproteusandTrypanosoma, although previous studies of blood parasites and predation also showed similar variation among taxa (Møller & Nielsen, 2007). We hypothesize that this effect was due to the impact of malaria as a cause of dramatic increases in the abundance of damaged erythrocytes, thereby severely reducing the efficiency of metabolism, but also restricting blood flow to the brain and the spleen (Atkinson & Van Riper, 1991; Chen et al, 2001). In addition blood parasites cause immune responses, fever and induction of heat-shock proteins (Wakelin, 1996; Merino et al, 1998; Chen et al, 2001), and these effects may affect the ability of an individual to escape when approached by a car. Such effects will be particularly strong for intra-erythrocytic blood parasites likePlasmodiumthat is the cause of severe anemia. Bird species with blood parasite infections have shorter flight distances than uninfected species (Møller, 2008b), suggesting that species with high level infections may take greater risks when encountering potential danger. The increase in mortality risk by traffic linked to blood parasite infections selects against blood parasite infections. The findings reported here also have implications for parasitological studies of blood parasites. Given that traffic-induced mortality disproportionately biases estimates of prevalence of blood parasites downwards, any epidemiological inference based on prevalence in populations of hosts is likely to be biased likewise.
Given the relationship between blood parasite infection and susceptibility to traffic, we should also expect immunity to explain interspecific variation in risk of being killed by a car. Indeed, we found a positive relationship between susceptibility to car accidents and size of the bursa of Fabricius. The bursa is an important immune defense organ involved in production of antibodies and responsible for differentiation of the entire repertoire of B-cells in young birds (Glick, 1983, 1994; Toivanen & Toivanen, 1987). This effect was independent of body mass.
Susceptibility to mortality caused by traffic increased with increasing body mass, when analyzing species-specific data and contrasts. Large bird species need longer distances for take-off, and climb rates decrease with body mass (Hedenström & Alerstam, 1992), and species with large body mass have long flight distances (Møller, 2008a,b). That may partly explain why species with large body mass run a higher risk of mortality due to traffic than small species, although other factors relating to body size may explain this variation. The best-fit model showed a negative relationship with body mass still being present after other predictor variables had been included.
We also investigated susceptibility to traffic in relation to habitat, flight behavior, duration of development and several other factors, but found little evidence of these factors explaining interspecific variation once other factors were included in the phylogenetic analyses.
In conclusion, we have found that risk of mortality in birds due to traffic increased with abundance. Once abundance had been taken into account, solitary species with short flight distances having a high prevalence of malarial parasites and a large bursa of Fabricius were particularly likely to die from traffic accidents. These findings suggest that risk of mortality due to traffic is associated with ability to escape an approaching vehicle. Furthermore, given that mortality due to traffic potentially is a non-negligible cause of mortality in birds the results suggest that car-induced mortality will select for resistance to blood parasites, increased sociality, and longer flight distances.
Anon. 2000. JMP.®[M]. Cary: SAS Institute Inc.
Atkinson C, Van Riper III C. 1991. Pathogenicity and epizootiology of avian haematozoa:Plasmodium,LeucocytozoonandHaemoproteus[M]//Loye JE, Zuk M (eds). Bird-Parasite Interactions. Oxford: Oxford University Press, 19-48.
Barker FK, Barrowclough GF, Groth JG. 2001. A phylogenetic hypothesis for passerine birds: Taxonomic and biogeographic implications of an analysis of nuclear DNA sequence data[J].Proc R Soc Lond B, 269: 295-308.
Batten LA. 1973. Population dynamics of suburban Blackbirds[J].Bird Study, 20: 251-258.
Bibby CJ, Hill DA, Burgess ND, Mustoe S. 2005. Bird Census Techniques[M]. London: Academic Press.
Blondel J, Catzeflis F, Perret P. 1996. Molecular phylogeny and the historical biogeography of the warblers of the genusSylvia(Aves) [J].J Evol Biol, 9: 871-891.
Blümel H, Blümel R. 1980. Wirbeltiere als Opfer des Strassenverkehrs[J].Abh Berliner Naturkundemus Görlitz, 54: 19-24.
Blumstein DT. 2006. Developing an evolutionary ecology of fear: How life history and natural history traits affect disturbance tolerance of birds[J].Anim Behav, 71: 389-399.
Bruun-Schmidt J. 1994. Trafikdræbte dyr i relation til landskab, topografi og vejtype[D]. M.Sc. thesis, Biological Institute, Odense University, Denmark.
Burnham KP, Anderson DR. 2002. Model Selection and Multi-model Inference[M]. New York: Springer.
Chen M, Shi L, Sullivan Jr D. 2001.HaemoproteusandSchistosomasynthesize heme polymers similar toPlasmodiumhemozoin and β-hematin[J].Mol Biochem Parasitol, 113: 1-8.
Cohen J. 1988. Statistical Power Analysis for the Behavioral Sciences[M]. 2nded. Hillsdale: Lawrence Erlbaum.
Cramp S, Perrins CM. 1977-1994. Handbook of the Biirds of Europe the Middle East and North Africa: The Birds of the Western Palearctic. Vols 1-9 [M]. Oxford: Oxford University Press.
Crochet PA, Bonhomme F, Lebreton JD. 2000. Molecular phylogeny and plumage evolution in gulls (Larini) [J].J Evol Biol, 13: 47-57.
Draper NR, Smith H. 1981. Applied regression analysis[M]. 2nded. New York: John Wiley.
Dunthorn AA, Errington FP. 1964. Casualties among birds along a selected road in Wiltshire[J].Bird Study, 11: 168-182.
Erritzøe J, Mazgajski TD, Rejt L. 2003. Bird casualties on European roads — A review[J].Acta Ornithol,38: 78-93.
Felsenstein J. 1985. Phylogenies and the comparative method[J].Am Nat, 125: 1-15.
Finnis RG. 1960. Road casualties among birds[J].Bird Study, 7: 21-32.
Fleishman LJ, Marshall CJ, Hertz PE. 1995. Comparative study of temporal response properties of the visual system of three species of anoline lizards[J].Copeia, 1995: 422-431.
Forman RTT, Alexander LE. 1998. Roads and their major ecological effects[J].Ann Rev Ecol Syst, 29: 207-231.
Garamszegi LZ, Erritzøe J, Møller AP. 2002. Coevolving eye size and brain size in relation to prey capture and nocturnality[J].Proc R Soc Lond B, 269: 961-967.
Garamszegi LZ, Erritzøe J, Møller AP. 2003. The evolution of immune defense and song complexity in birds[J].Evolution, 57: 905-912.
Garamszegi LZ, Erritzøe J, Møller AP. 2007. Feeding innovations and immune defense in birds[J].Biol J Linn Soc, 90: 441-455.
Garamszegi LZ, Møller AP. 2007. Prevalence of avian influenza and host ecology[J].Proc R Soc Lond B, 274: 2003-2012.
Garland Jr T, Harvey PH, Ives AR. 1992. Procedures for the analysis of comparative data using phylogenetically independent contrasts[J].Syst Biol, 41: 18-32.
Glick B. 1983. Bursa of Fabricius: vol.7[M]// Farner DS, King JR. Avian Biology. New York: Academic Press. 443-500.
Glick B. 1994. The bursa of Fabricius: The evolution of a discovery[J].Poult Sci, 73: 979-983.
Gliwicz J, Goszczynski J, Luniak M. 1994. Characteristic features of animal populations under synurbanization — the case of the blackbird and the striped field mouse[J].Mem Zool, 49: 237-244.
Göransson G, Karlsson J, Lindgren A. 1978. Vägars Inverkan på Omgivande Natur. II. Fauna. (Impact of roads on surrounding environment) [M] Stockholm: SNV Forskningsnämnden.
Grell MB. 1998. Fuglenes Danmark[M]. Copenhagen: Gad.
Haas W. 1964. Verluste von Vögeln und Säugern auf Autostrassen[J].Ornithol Mitt, 16: 245-250.
Hansen L. 1969. Trafikdøden i den danske dyreverden[J].Dansk Ornithol Foren Tidsskr, 63: 81-92.
Hansen L. 1982. Trafikdræbte dyr[J].Dansk Ornithol Foren Tidsskr, 76: 97-110.
Hedenström A, Alerstam T. 1992. Climbing performance of migrating birds as a basis for estimating limits for fuel-carrying capacity and muscle work[J].J Theor Biol, 164: 19-38.
Heinrich D. 1978. Untersuchungen zur Verkehrsopferrate bei Säugetieren und Vögeln[J].Zeitschr Natur- Landeskde Schleswig-Holstein Hamburg, 85: 193-208.
Helbig AJ, Seibold I. 1999. Molecular phylogeny of Palearctic-AfricanAcrocephalusandHippolais(Aves: Sylviidae)[J].Mol Phylogenet Evol, 11: 246-260.
Hodson NL. 1960. A survey of vertebrate road mortality 1959[J].Bird Study, 7: 224-231.
Hodson NL, Snow DW. 1965. The road deaths enquiry, 1960-61[J].Bird Study, 12: 90-99.
Jones KE, Purvis A. 1997. An optimum body size for mammals? Comparative evidence from bats[J].Funct Ecol, 11: 751-756.
Klausnitzer B. 1989. Verstädterung von Tieren[M]. Wittenberg-Lutherstadt: Neue Brehm-Bücherei.
Maddison WP, Maddison DR. 1992. MacClade[M]. Version 3.0. Sunderland: Sinauer.
Merino S, Martínez J, Møller AP, Barbosa A, de Lope F, Rodriguez-Caabeiro F. 1998. Blood stress protein levels in relation to sex and parasitism of barn swallows (Hirundo rustica) [J].Ecoscience, 9: 300-305.
Møller AP. 2008a. Flight distance of urban birds, predation and selection for urban life[J].Behav Ecol Sociobiol, 63: 63-75.
Møller AP. 2008b. Flight distance and blood parasites in birds[J].Behav Ecol, 19: 1305-1313.
Møller AP. 2009. Successful city dwellers: A comparative study of the ecological characteristics of urban birds in the Western Palearctic[J].Oecologia, 159: 849-858.
Møller AP, Birkhead TR. 1994. The evolution of plumage brightness in birds is related to extra-pair paternity[J].Evolution, 48: 1089-1100.
Møller AP, Erritzøe J. 1996. Parasite virulence and host immune defence: Host immune response is related to nest re-use in birds[J].Evolution, 50: 2066-2072.
Møller AP, Erritzøe J, Garamszegi LZ. 2005. Coevolution between brain size and immunity in birds: Implications for brain size evolution[J].J Evol Biol, 18: 223-237.
Møller AP, Erritzøe J, Nielsen JT. 2010. Causes of interspecific variation in susceptibility to cat predation on birds[J].Chn Birds, 1: 97-111.
Møller AP, Nielsen JT. 2007. Malaria and risk of predation: A comparative study of birds[J].Ecology, 88: 871-881.
Møller AP, Nielsen JT, Garamszegi LZ. 2008. Risk taking by singing males[J].Behav Ecol, 19: 41-53.
Nankinov DN, Todorov NM. 1983. Bird casualties on highways[J].Soviet J Ecol, 14: 288-293.
Neter J, Kutner MH, Nachtsheim CJ, Wasserman W. 1996. Applied Linear Statistical Models[M]. Chicago: Irwin.
Purvis A, Rambaut A. 1995. Comparative analysis by independent contrasts (CAIC) [J].Comp Appl Biosci, 11: 247-251.
Reijnen R, Foppen R. 1991. Effect of road traffic on the breeding sitetenacity of male willow warblers (Phylloscopus trochilus) [J].J Ornithol, 132: 291-295.
Reindt FE. 2003. The impact of roads on birds: Does song frequency play a role in determining susceptibility to noise pollution? [J].J Ornithol, 144: 295-306.
Roberts SC. 1988. Social influence on vigilance in rabbits[J].Anim Behav. 36: 905-913.
Rosenthal R. 1991. Meta-analytic Procedures for Social Research[M]. New York: Sage.
Scheuerlein A, Ricklefs RE. 2004. Prevalence of blood parasites in European passeriform birds[J].Proc R Soc Lond B, 271: 1363-1370.
Shochat E, Warren PS, Faeth SH, McIntyre NE. 2006. From patterns to emerging processes in mechanistic urban ecology[J].Trends Ecol Evol, 21: 186-191.
Sibley CG, Ahlquist JE. 1990. Phylogeny and Classification of Birds, a Study in Molecular Evolution[M]. New Haven: Yale University Press.
Smettan HW. 1988. Wirbeltiere und Strassenverkehr - ein ökologischer Beitrag zum Strassentod von Säugern und Vögeln am Beispiel von Ostfildern/Württemberg[J].Ornithol Jahrb Baden-Württemberg, 4: 29-55.
Sokal RR, Rohlf FJ. 1995. Biometry[M]. 3rded. New York: Freeman.
Starck JM, Ricklefs RE. 1998. Avian Growth and Development. Evolution within the Altricial-precocial Spectrum[M]. New York: Oxford University Press.
Stephan B. 1999. Die Amsel[M]. Wittenberg-Lutherstadt: Neue Brehm-Bücherei.
Straube H. 1988. Tiere am Strassenrand[J].Collurio, 1988: 28-39.
Svensson L. 2006. Identification Guide to European Passerines[M]. Thetford: British Trust for Ornithology.
Svensson S. 1998. Bird kills on roads: is this mortality factor seriously underestimated? [J].Ornis Svecica, 8: 183-187.
Toivanen P, Toivanen A. 1987. Avian Immunology: Basis and Practice[M]. Boca Raton: CRC Press.
Underhill JE, Angold PG. 2000. Effects of roads on wildlife in an intensively modified landscape[J].Environ Rev, 8: 21-39.
Vignes JC. 1984. Les oiseaux victimes de la circulation routière au Pays basque français[J].Oiseau, 54: 137-148.
Wakelin D. 1996. Immunity to Parasites: How Parasitic Infections Are Controlled[M]. Cambridge: Cambridge University Press.
Warren PS, Katti M, Ermann M, Brazel A. 2006. Urban bioacoustics: It's not just noise[J].Anim Behav, 71: 491-502.
Wäscher S, Janisch A, Sattler M. 1988. Verkehrsstrassen - Todesfallen der Avifauna[J].Luscinia, 46: 41-55.
鸟类交通致死原因的种间行为差异分析
Anders Pape Møller1,*, Helga Erritzøe2, Johannes Erritzøe2
(1.Department of Ecology, Systematics and Evolution, CNRS UMR8079,University of Paris-South, Bâtiment362,F-91405Orsay Cedex,France; 2.Taps Old Rectory, DK-6070Christiansfeld,Denmark)
鸟类和其他动物经常死于车撞, 每年因车撞而死亡的个体可达数百万只。为什么有的物种比其他物种更易于死于车撞的问题,迄今未研究过。该文推测物种本身所具有的冒险行为可能是这些物种的某些个体更常死于车撞的一个原因。此外,不同物种个体在公路上的行为、个体数量、栖息地偏好、繁殖社会性以及健康状况都可能是影响个体车撞风险的潜在因素。2001—2006年,对丹麦一个农村地区进行定期统计取样, 得到该地区不同种鸟类个体被车撞死事件发生的频率,以及周边环境中 50种鸟类的个体数量数据, 并利用这些数据来检验我们的预测。车撞频率随鸟类个体数量线性增加和位于道路上或者低空飞越道路的个体所占比例增加,这两个因素几乎无法解释鸟类车撞频率差异。在考虑到丰富度这个因素后,发现那些鸟类惊飞距离短、面对潜在危险而更具有冒险性的物种个体更易遭遇车撞而亡。另外, 独居的物种、疟原虫感染率高的物种及因体型大而拥有大法氏囊的物种个体死于车撞的频率更高。这些发现表明, 一系列反映冒险行为、视敏度和健康状况的因子导致某些鸟类物种易受到车撞影响。
鸟类;血液寄生虫; 惊飞行为;惊飞距离; 栖息地选择;社会性
Q958.11;Q959.7
A
0254-5853-(2011)02-0115-13
2010-06-17;接受日期:2011-02-23
10.3724/SP.J.1141.2011.02115
date: 2010-06-17; Accepted date: 2011-02-23
*Corresponding author (通讯作者), Email: anders.moller@u-psud.fr
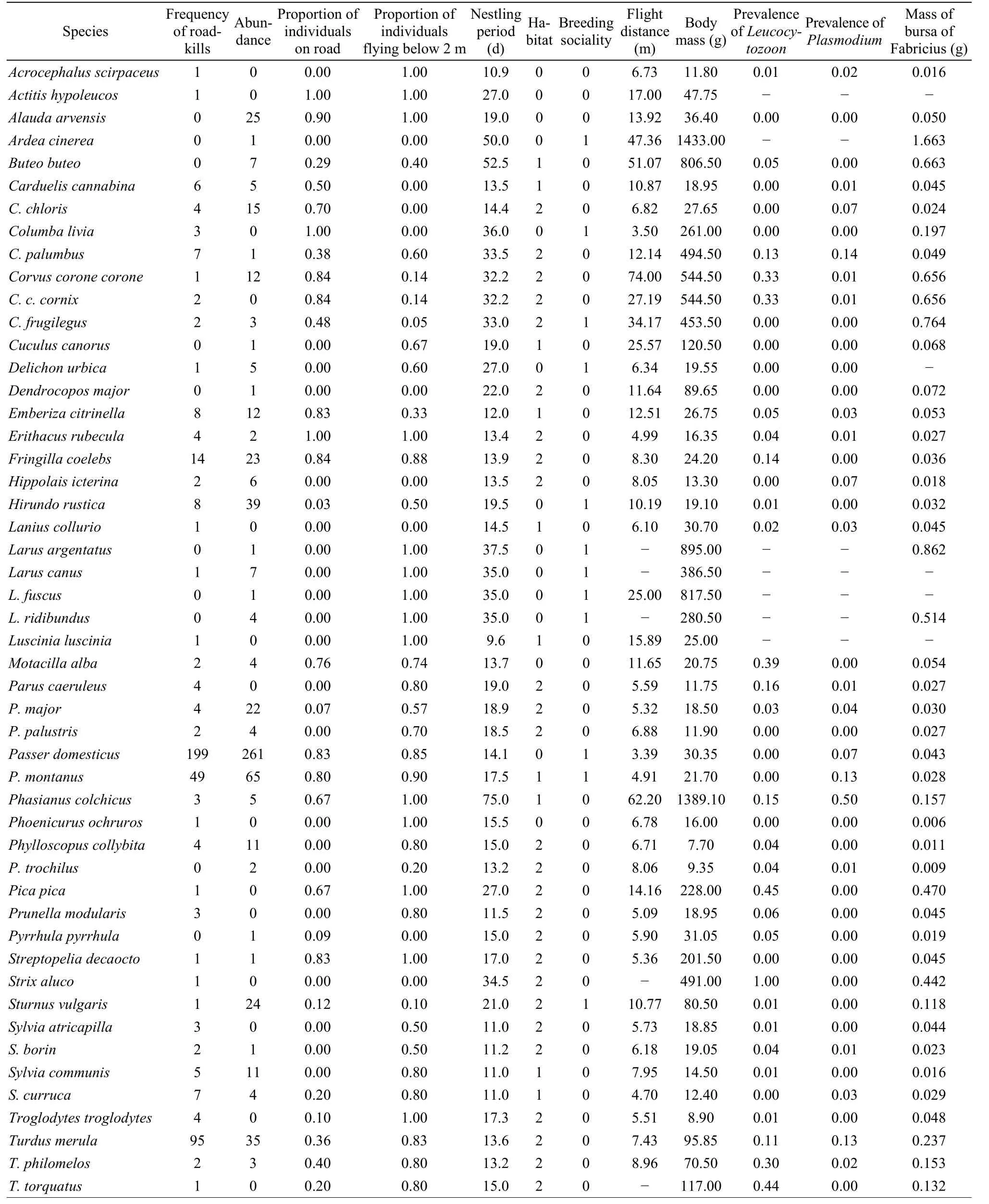
Appendix 1 Frequency of road-kills of 50 species of birds, abundance of these species during standard point count censuses, proportion of individuals recorded on the road, proportion of individuals flying below 2 m, duration of nestling period (days), habitat (0 — grassland, 1 —bushes, 2—trees), breeding sociality (0 — solitary, 1 — colonial), mean flight distance (m), body mass (g), prevalence of Leucocytozoon and Plasmodium, and mass of bursa of Fabricius (g)
