低能离子注入彩棉种子的生物物理机理
2011-12-18刘效勇黄旭初
孙 毅 刘效勇 井 群 黄旭初
(1.昌吉学院物理系 新疆 昌吉 831100; 2.石河子大学生态物理重点实验室 新疆 石河子 832000; 3.石河子大学师范学院物理系 新疆 石河子 832000;)
低能离子注入彩棉种子的生物物理机理
孙 毅1刘效勇2,3井 群2,3黄旭初2,3
(1.昌吉学院物理系 新疆 昌吉 831100; 2.石河子大学生态物理重点实验室 新疆 石河子 832000; 3.石河子大学师范学院物理系 新疆 石河子 832000;)
通过建立与生物组织较为接近的靶模型,采用MonteCarlo方法研究了低能离子注入植物种子诱变的机理,为进一步指导育种实践提供重要的理论指导。结果表明在 20-200keVN+注入彩棉种子,离子的深度在 0-22μm之间,对于单个细胞大量离子被阻停在细胞质内较浅的区域,这段区域很可能是低能离子与生物组织发生三因子效应的主要区域,对于“马鞍型”曲线产生的原因推测是由能量、质量和深度等因素共同决定。
离子注入;蒙特卡罗模拟;种子;存活剂量关系
离子束用于诱变育种、细胞加工、转基因等方面,已经取得显著的进展[1,2]。但是,到目前为止,这方面的理论研究尚很不完善,尤其是低能离子束生物效应的机理仍然很不清楚。问题的关键是质量、能量、电荷三位一体的低能离子怎样进入生物体,进入生物体后沉积的深度 -浓度分布规律,以及沉积所造成的影响[3]。基于生物组织内含有 H、N等元素,造成实验上检测 H+、N+等与生物体的相互作用有一定的困难,现有的理论计算方法多局限于重离子与生物组织的相互作用[4],且计算方法有一定的局限性[5],为此本文提出一个较接近于实际的生物靶模型和相对可靠的计算方法,为研究低能离子束与生物体之间的原初作用提供了理论依据。
1 物理模型
将植物种子中的单个细胞划分成细胞壁、细胞膜、细胞质、亚细胞颗粒四部分。用纤维素 (C6H10O5)n来模拟细胞壁,膜磷脂NC41H80O8P近似细胞膜的主要成份,水 H2O近似细胞质,假设各种亚细胞颗粒(随机细胞器)分布在细胞质内,亚细胞颗粒的大小一般在 1-5μm之间,其各元素的原子比可以通过靶材料内蛋白质和脂肪的组成与含量得到。试验测得彩棉种子的组成与含量分别为:水分 6.1%、脂肪 33.4%、蛋白质 38.5%、碳水化合物 14.6%、纤维素 2.1%和无机盐 5.3%。根据蛋白质与脂肪的组成和含量,可以计算出亚细胞颗粒的分子组成近似为(忽略碳水化合物、无机盐)C16H30N3O5。根据细胞结构建立的靶模型参数见表1 ,如果离子能够穿过这 4个区域将进入下一个循环。

表1 模型的主要参数Table1 Main parametes of the model
2 计算方法
2.1 核碰撞能量损失根据经典的两体碰撞理论,粒子同靶原子的单次散射后能量转移为:

其中M1、M2分别为入射离子和靶原子质量,E0为入射粒子的初始能量,θc为质心系下的入射粒子的散射角。若散射截面为 dσ=2πpdp,其中 p为碰撞参数,靶材料的原子密度为 N,那么当该粒子在固体中穿行长度为Δx时,可以得到核阻止本领的表示式:

其中 Sn表示核阻止截面,由下式给出:

2.2 射程的计算
近似认为核碰撞能量损失和电子碰撞能量损失相互独立,在电子碰撞能量损失忽略的情况下,入射粒子在单位路径上的总能量损失为:

根据布拉格提出的阻止截面具有线性叠加的特性,对于具有相当于AmBn组成的混合物其阻止截面由下式给出:

其中,SA和 SB分别是A原子和B原子的阻止截面。分别计算 C、H、O、N元素总的核阻止截面 Sn,可得到入射粒子在生物材料中的总阻止截面 S。若用 R表示粒子在靶材料中的实际穿行路程,那么粒子在靶材料中的总射程可以表示为:

2.3 碰撞参数和自由程的确定
在低能情况下,碰撞自由程L(N)可以近似为原子间的平均间距,若每个单位步长距离 Si是随机的,则直接抽样可得碰撞步长,表示为:Si=-L(N)lnξ1,其中N表示靶原子密度,ξ1表示[0,1]之间的随机数。在程序设计中,通过长度为L(N)的飞行路程中随机抽样得到可能发生的碰撞的碰撞参数 p= [-lnξ2/πNL(N)]1/2,其中ξ2表示[0,1]之间的随机数。
2.4 散射角和方位角的确定
入射离子在靶材料中的散射角与微分散射截面 dσ有关,由碰撞参数通过数值计算可以得到相应的散射角,但散射角应满足一定的范围[θmin,π],其中θmin的大小由下式决定:

方位角Ψ为 0到 2π之间随机分布,因此可以抽样得到:Ψ =2π ξ3,其中ξ3为 0到 1之间的随机数。
3 计算结果与分析
根据上述原理,采用MonteCarlo方法模拟计算N+注入彩棉种子,近似认为注入离子与靶原子在经过自由程碰撞后平均偏转角小于 5度,当注入离子能量小于 5eV终止循环。通过跟踪 5000个离子历史,分别记录能量损失和射程。
3.1 能量与射程分布
根据带电粒子在靶材料中的能量损失机制,在 20-200keV能量区间电子阻止可以忽略,只计算核碰撞能量损失,通过计算得到靶模型 4个区域内阻止本领和能量之间的关系 (图 1)。从图 1可以看出在 4个不同区域,核阻止本领和能量的变化关系略有不同。细胞质区间曲线变化较为平缓,其他 3个区间随着能量的增加核阻止本领在逐渐减弱,在 20-80keV区间去曲线相对较陡,在 80-200keV区间曲线相对平缓。离子在细胞壁、细胞膜和随机细胞器中的核阻止与电子阻止随能量的变化关系相差不大,在细胞质区域核阻止相对较小,这与不同区域内靶材料的组成相关。由于随机细胞器主要由蛋白质和脂肪组成,在很小的体积内蛋白质结构近似于晶体结构,因此在这个区域内对离子的射程影响最为明显。然而在细胞质内是由水构成的,离子在这个区域内受到的阻止作用就相对小一些。相关文献计算表明,当能量在 103(或更小一些)-105eV范围时,核阻止本领近似为与离子能量无关的常数[6,7]。在本文的物理模型下,计算表明只有在细胞质区域核阻止本领才可以近似为与离子能量无关的常数,这说明在计算离子束与生物组织相互作用过程中使用这个近似关系,即用水来近似生物组织的主要成分。在能量为 20-80keV之间曲线变化明显,这表明随着入射深度的增加,离子能量减少,核阻止本领增加,当能量减少到对应于核阻止本领 -能量曲线起始位置时,能量损失急剧增加,其剩余能量在很短的射程内几乎全部损失掉,从而会形成一个能量沉积的Bragg峰。
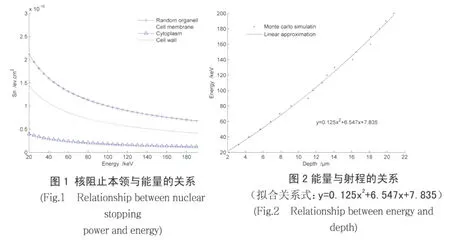
通过计算总阻止本领和能量的关系,代入射程的计算式可以得到能量与射程的关系(图 2)。从图 2可以看出在 20-200keV区间,射程为 0-22μm区间,N+注入彩棉种子的射程随能量的增加逐渐增加,经过拟合可以得到一个较为简单的多项式:y=0.125x2+6.547x+7.835。实验上通常选择的能量在 20 -40keV之间,由计算得到的相应射程为 2-4.5μm,在细胞质的浅层区域,表明这段区域很可能是低能离子与生物组织发生三因子效应(质量沉积、能量沉积、电和交换)的主要区域。
3.2 能量沉积作用
图 3为 30keVN+在靶模型中 4个不同区域内的能量沉积曲线。从图 3可以看出 4条曲线的总体趋势一致,曲线前端较为平缓,末端急剧上升,其能量主要沉积在细胞的浅层区域,说明低能离子对细胞浅层区域的微观结构损伤最大,对细胞表面性能的损伤也最厉害。对于单个细胞,细胞壁和细胞膜占整个细胞的比例较少,能量沉积可以忽略不计。随着入射深度的增加离子能量沉积也逐渐增加,细胞质和随机细胞器 2个不同区域能量沉积分别在 1.96μm和 2.62μm处达到最大值。

3.3 离子浓度与生物效应
实验上主要通过测量重离子的浓度 -深度分布来分析离子束与生物组织的原初作用,考虑到不同离子所产生的生物效应有较大的差异,在本文的物理模型下计算了 30keVN+注入彩棉种子的浓度 -深度分布,计算结果见图 4。从图 4可以看出 N+离子的浓度 -深度曲线呈现为有拖尾的非对称高斯分布,峰值位置在 2.50μm,大量离子被阻停在 0-10μm的深度范围。根据假设模型在这个区域仍然在单个细胞内,大量离子穿过细胞膜和细胞壁,停留在细胞质的浅层,部分与随机细胞器发生作用。文献[5]采用 Tr im程序计算得到能量碰撞损失,然后根据实验结果对 LSS理论修正得到 20keVN+注入棉花种子的最可几深度(离子浓度最大值所对应的深度)是 8μm,最大深度是 20μm,本文认为这个计算结果有待考究。基于实验手段和仪器的限制,目前无法从实验上测量N+注入棉花种子的射程分布,这就使得无法根据实验结果对LSS理论中的能量损失进行修正,而由 Trim程序计算得到的能量损失与实验结果有较大的差异,不能用于计算低能离子与生物组织之间的相互作用[8]。本文的计算在对靶材料进行简化改进的基础上,通过经典的两体碰撞理论和严格的数值计算,没有使用相关的经验公式,与对 LSS理论进行修正的计算方法相比是一种相对可靠的计算方法。

在离子注入生物学效应研究中,N+注入的剂量 -效应关系呈特有的类“马鞍型”,对此类曲线产生的机理有质量沉积模型、自由修复模型和碎片重组修复模型等多种不同的解释[10]。对于这些解释都与低能离子与生物组织的原初作用相关,即在深度小的区域生物效应弱,在深度大的区域生物效应强,这暗示着深度和生物效应之间存有一定联系。根据本文的物理模型若将相对离子浓度和能量作为物理参量,将离子的注入深度转化为相应的生物效应,发现经过叠加所得到的参量 -效应关系曲线与剂量 -效应关系曲线有很高的相似性(见图 5)。从图中可以看到 2条趋向都呈现为先下降再升高的“马鞍型”,但曲线的最低转折点和鞍点的位置有所不同。由此推测“马鞍型”曲线产生的原因:在特定的能量下,剂量与效应之间的关系可以认为是由能量和浓度 2个主要参量与深度之间关系的叠加,认为是能量沉积和质量沉积两方面因素所导致。基于参量 -效应关系曲线是在假设深度和效应之间有一定的联系的基础上得到,要对“马鞍型”曲线作出更有说服力的理论解释,还需要更多的实验依据。
4 结语
通过理论计算表明 20-200keV的N+在彩棉种子中的深度随着能量的增加逐渐增加,它们之间可以表示为一个简单的多项式形式,在此能量区间离子的注入深度为 0-22μm,对于单个细胞大部分离子停留在细胞质区域,能量沉积也主要发生在细胞质和随机细胞器区域,在不考虑 X射线、电荷交换及其他一些次级作用的基础上,离子不太可能穿过单个细胞。低能离子对植物种子有明显的诱变作用[11],这说明仅用核碰撞能量损失来解释离子束与生物体之间的相互作用是不合适的,电荷交换及碰撞产生的生物效应可能是诱变的主要原因。最近等离子体所离子束生物工程学院重点实验室在前期辐射远程效应研究的基础上,进一步证实在植物个体水平辐射诱导的远程诱变效应的存在,为低能离子的诱变机理提供了重要的实验依据。低能离子束诱变机理的研究在探索生命过程和定向改良品种两方面都将发挥越来越重要的作用。
[1]余增亮,邱励俭,霍裕平.离子注入生物效应及育种研究进展[J].安徽农学院学报,1991,18(4):251-257.
[2]曾宪贤,武宝山,吕杰.离子束生物技术在生命科学中的应用[J].核技术,2006,29(2):112-115.
[3]卫增泉,韩光武,周光明,李强,颉红梅.超低能离子注入作物育种的一种重要机制[J].生物物理学报,1996,12 (2):315-320.
[4]张丰收.离子与生物分子相互作用的微观动力学理论[J].原子核物理评论,2006,23(1):23-43.
[5]王林香,祝恒江,王世亨,王先明.低能 N+注入植物种子射程分布的模拟计算[J].高能物理和核物理,2007,31 (3):311-315.
[6]吕晓桂,罗辽复.低能离子束在植物种粒和微生物中的穿透深度[J].生物物理学报,2006,22(1):57-61.
[7]余增亮.离子束生物技术引论[M].合肥:安徽科学技术出版社,1998:41-45.
[8]汪新福,陆挺,周宏余,朱光华,丁晓纪.低能 V+注入花生种子的深度研究[J].原子能科学技术,2002,36(6): 531-534.
[9]王新绘,李冠,吕杰,张秋云.氮离子注入彩棉种子引起M1代的变化[J].激光生物学报,2004,13(4):294-297.
[10]李珂,张东正,赵瑾,张根发.重离子束注入与生物体的相互作用及遗传诱变的分子机制[J].原子核物理评论, 2007,24(2):117-123.
[11]马飞,冯伟森,张莉,王从晓,张学品,吴少辉,温红霞,高海涛.离子束生物技术在药用植物遗传改良中的应用前景[J].中国农学通报,2010,26(4):37-40.
Q319.2
A
1671-6469(2011)02-0099-06
2011-02-24
昌吉学院自然科学基金项目资助(08YJYB012)
孙毅 (1981-),男,新疆喀什人,昌吉学院物理系,讲师,在读博士,研究方向:凝聚态物理研究。
(责任编辑:代琴)
