转cry1Ab基因粳稻对稻田节肢动物群落的影响
2011-11-21刘志诚田俊策卢增斌舒庆尧彭于发叶恭银
刘志诚, 陈 洋, 田俊策, 卢增斌, 舒庆尧, 胡 萃, 彭于发, 叶恭银
1浙江大学昆虫科学研究所,水稻生物学国家重点实验室,农业部作物病虫分子生物学重点开放实验室;2浙江大学原子核农业科学研究所,浙江 杭州310029;3中国农业科学院植物保护研究所,植物病虫生物学国家重点实验室,北京100193
转cry1Ab基因粳稻对稻田节肢动物群落的影响
刘志诚1, 陈 洋1, 田俊策1, 卢增斌1, 舒庆尧2, 胡 萃1, 彭于发3, 叶恭银1
1浙江大学昆虫科学研究所,水稻生物学国家重点实验室,农业部作物病虫分子生物学重点开放实验室;2浙江大学原子核农业科学研究所,浙江 杭州310029;3中国农业科学院植物保护研究所,植物病虫生物学国家重点实验室,北京100193
在将稻田节肢动物群落按营养关系分为植食类、寄生类、捕食类、腐食类和其他类等5个功能团的基础上,从功能团优势度、群落结构参数及群落相异性等方面,经2年3点的调查就2个转cry1Ab基因粳稻(Bt粳稻)品系KMD1和KMD2对稻田节肢动物群落结构的影响做了评价。结果表明:在大多数情况下,Bt粳稻与对照间各功能团优势度、群落结构参数[物种丰富度(S)、Shannon-Wiener多样性指数(H′)、均匀性指数(J)、优势集中性指数(C)]及其时间动态无明显差异;Bt粳稻与对照间植食类、寄生类、捕食类亚群落,及整个节肢动物群落的相似性也较高。综合分析认为,Bt粳稻对稻田节肢动物群落结构无明显的负面影响。
Bt粳稻;cry1Ab基因; 节肢动物; 功能团; 群落结构
随着转Bt基因抗虫棉花等的商业化应用,截至2009年全球种植转基因抗虫作物的面积已达2170万hm2(James,2010),人们也越来越关注转基因作物种植的生态安全性问题,这些问题主要涉及转基因抗虫作物对作物生态系统生物多样性、生物群落结构的影响等。目前,关于Bt棉花(Dhawanetal.,2009; Headetal.,2005; Sistersonetal.,2004; Whitehouseetal.,2005; 崔金杰和夏敬源,2000) 、Bt玉米(Bitzeretal.,2005; Obryckietal.,2001)、Bt马铃薯(Riddick amp; Barbosa,1998)的安全性有些探讨,但大多数是评价转基因抗虫作物对几种主要天敌生物学特性或种群数量的影响,而缺乏对群落水平的全面系统评价。就Bt水稻而言,在决策是否商业化应用前及时评估其生态安全性颇为必要。迄今,一些培育得到的抗虫Bt水稻表现出对螟虫良好的抗虫性(Chenetal.,2010; Dattaetal.,1998; Shuetal.,2000; Tuetal.,2000; Yeetal.,2001、2003),但在其生态安全性评价方面,仅就Bt水稻对非靶标害虫(Akhtaretal.,2010; Chenetal.,2007)和蜘蛛(Tianetal.,2010; 刘志诚等,2002)的优势种田间种群动态的影响做了评价,尚需进行更全面的研究。
稻田节肢动物群落是一个以水稻为中心,由多种植食性昆虫、天敌(如寄生性昆虫、捕食性昆虫和蜘蛛等)、腐食性昆虫和其他节肢动物通过食物链相互联系而组成的复杂网络系统。稻田节肢动物的种类和数量相当丰富,若将整个节肢动物群落划分为不同的功能团,并以此来探讨群落的结构和多样性变化,即可使复杂的网络结构简单化,不同类群间的关系更加明晰(刘雨芳等,1999)。因此,有关稻田节肢动物群落结构的定量研究,多按照分类单元(如科、属等)或生活习性将节肢动物归类成不同功能团而进行(Heongetal.,1991; Schoenlyetal.,1998; 郝树广等,1998)。在这种系统中,任何营养层的某个物种生物量或物种组成的变化,都有可能通过食物链的营养流或化学信息流对其他营养层产生直接或间接的影响。因此,当Bt水稻种植于稻田生态系统后,因其对第二营养层螟虫(靶标害虫)等植食性鳞翅目昆虫的高致死作用,以及其本身物理性状、农艺性状、营养物质、挥发性和非挥发性次生化合物可能发生的非预期变化,极有可能对有关营养层的节肢动物种类组成、数量与发生动态等产生不同程度的影响,进而引起稻田生态系统中节肢动物群落结构发生变化。但事实究竟如何,目前尚难定论。为此,我们就Bt粳稻在不同稻田生态系统中对节肢动物群落结构的影响做了2年3点的调查与研究。
1 材料与方法
1.1 供试水稻
转基因抗虫Bt粳稻为纯合品系的“克螟稻1号(KMD1)”和“克螟稻2号(KMD2)”,其中,2000和2001年所用材料分别为R8、R9代。它们均采用农杆菌介导法获得,含cry1Ab基因和ubiquitin启动子基因,室内和田间条件下对螟虫等8种鳞翅目害虫表现高抗(Shuetal.,2000; Yeetal.,2001、2003)。对照为未转基因的亲本粳稻品种秀水11(CK)。
1.2 试验设计与调查方法
2000~2001年在浙江省选取3个试验点。按转基因作物安全管理要求,试验圃四周设置溪沟、蔬菜田或生育期与Bt粳稻不一致的水稻田作为自然隔离带。浙江大学实验农场水稻区和蔬菜区(无水稻种植史)设杭州点I和II,试验连续进行2年,同一田块中分设3个小区,每个材料为1个小区,每个小区面积为333 m2。对照区设于KMD1和KMD2小区间。2000年在安吉设置一点,就KMD1和对照做比较,两者小区面积均为1000 m2。各点的各小区分别单本插种各自的供试水稻材料。肥水管理按常规操作,水稻全生育期不施任何农药。移栽后30 d开始,每15 d取样调查1次,至水稻黄熟收割前10 d。每区采用对角线法选取5点,先用采样框(0.5 m×0. 5 m×0.9 m)罩住稻株,再用参照刘雨芳等(1999)改装的吸虫器采集取样。取样后立即用75%酒精保存,携回室内,除去杂物,挑出节肢动物,再用80%酒精保存。最后在体视镜下进行科、种的分类鉴定和计数。常见种鉴定至种,其他鉴定至科。
1.3 功能团的划分与群落多样性分析
参照郝树广等(1998)、Heongetal.(1991)和Schoenlyetal.(1998)将稻田节肢动物按营养关系划分为植食类、寄生类、捕食类、腐食类、其他类等5个功能团,计算并分析各功能团在调查所得节肢动物个体总数中的优势度。
优势度/%=Ni/N×100
式中,Ni为群落中第i个物种(科)的个体总数量,N为全部种(科)的个体总数量。

以科为单位,分别计算Bt粳稻和对照稻田节肢动物群落结构的有关参数,并比较分析Bt粳稻与对照间群落结构的相异性。有关计算公式如下:



式中,ni为第i个优势类群在群落中的重要值;N为群落的总重要值。
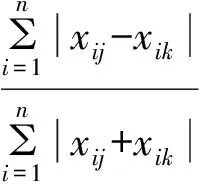
式中,n为样本中的物种数量;xij、xik分别为样本j和样本k中第i个类群的个体数。
各参数的计算用DPS(唐启义和冯明光,1997)软件进行。
2 结果与分析
2.1 功能团组成及其优势度
经2年3点调查,共获节肢动物个体37054头,隶属12目81科,归属5个功能团。其中,植食类包括7目27科昆虫,常见种有白背飞虱Sogatellafurcifera、褐飞虱Nilaparvatalugens、黑尾叶蝉Nephotettixcincticeps、稻纵卷叶螟Cnaphalocrocismedinalis和二化螟Chilosuppressalis;寄生类包括2目16科昆虫,常见种为茧蜂和金小蜂;捕食类包括7目27科昆虫或蜘蛛,常见种有拟水狼蛛Piratasubpiraticus、食虫沟瘤蛛Ummeliatainsecticeps、锥腹肖蛸Tetragnathamaxillosa和黑肩绿盲蝽Cyrtorrhinuslividipnnis等;腐食类共有3目8科昆虫,以鳞跳虫、圆跳虫或蠓为主;其他类共有1目5科,常见种为蚊和摇蚊。
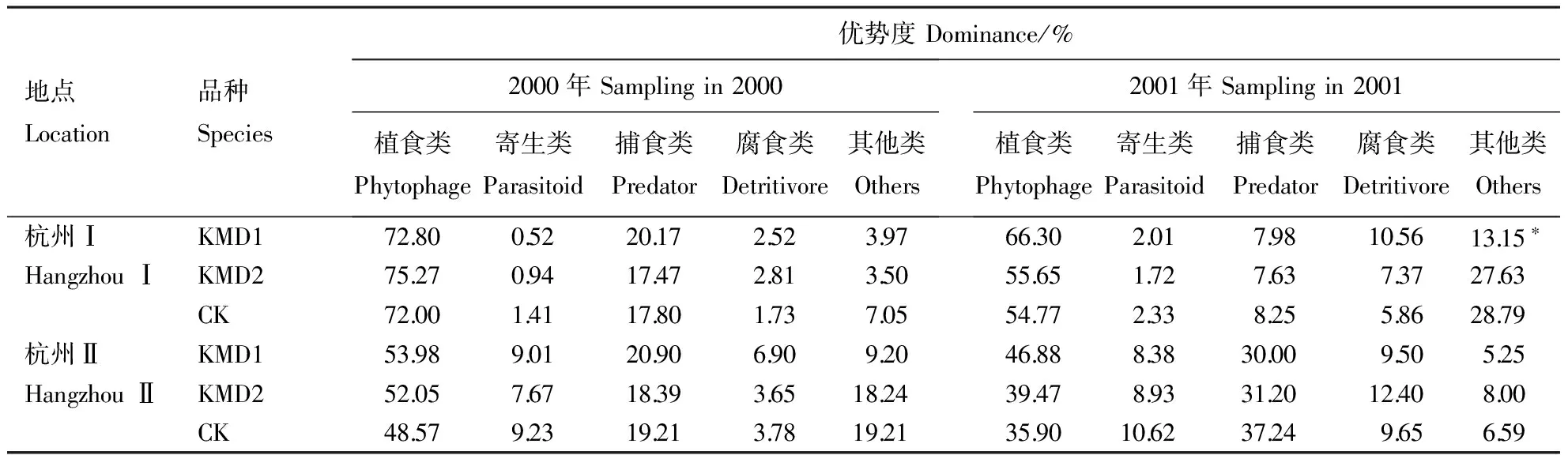
表1 转cry1Ab基因粳稻与对照间节肢动物功能团优势度的比较
2.2 群落结构参数及其时间动态
在杭州点,Bt粳稻与对照的节肢动物群落结构参数(S、H′、J、C)如表2所示。在安吉点,KMD1的各参数值分别为52、4.27、0.75和0.08,对照为55、4.41、0.76和0.07。可见,Bt粳稻与对照的群落结构参数基本接近,有的甚至相同。
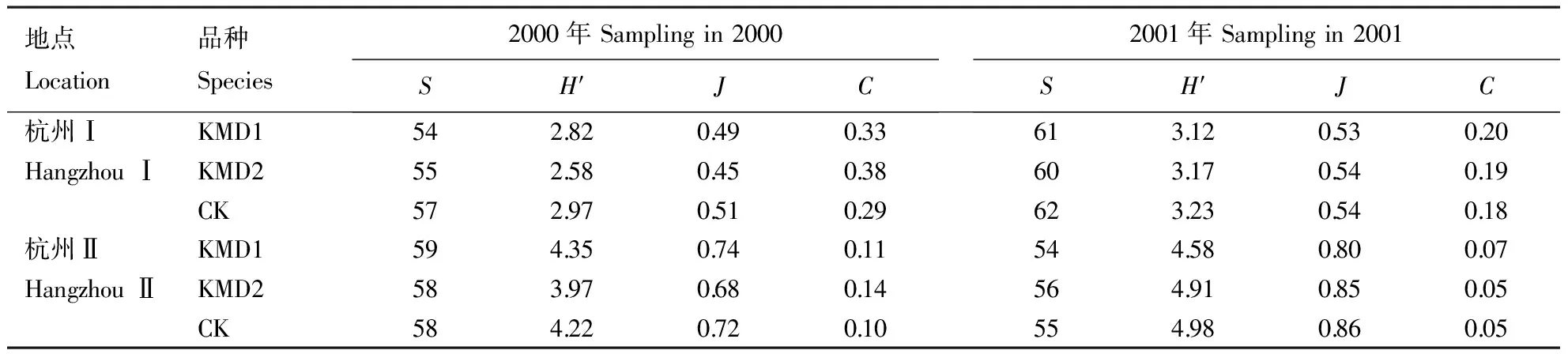
表2 转cry1Ab基因粳稻与对照间节肢动物群落结构参数的比较
就S、H′、J、C的时间动态趋势而言,Bt粳稻与对照间除个别时间略有差异外,大多数没有差别(图1~2)。
2.3 节肢动物群落相似性
在杭州点,Bt粳稻与对照间节肢动物群落的Bray-Curtis距离系数如表3所示。在安吉点,KMD1的植食类、寄生类、捕食类亚群落,及整个节肢动物群落与对照相应的亚群落或群落间的Bray-Curtis距离系数分别为0.19、0.49、0.12、0.15。综合分析可知,Bt粳稻与对照间植食类、寄生类、捕食类亚群落和整个节肢动物群落的相异性指数范围分别为0.09~0.29、0.11~0.61、0.12~0.38和0.09~0.30。除2000年杭州Ⅰ和安吉点的Bt粳稻的寄生类亚群落与对照间相异性较高外,其他均较低,即两者间相似性较高。
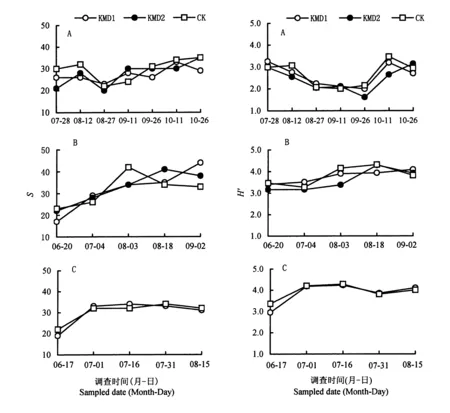

A: 杭州ⅠHangzhouⅠ; B: 杭州ⅡHangzhouⅡ; C: 安吉 Anji。

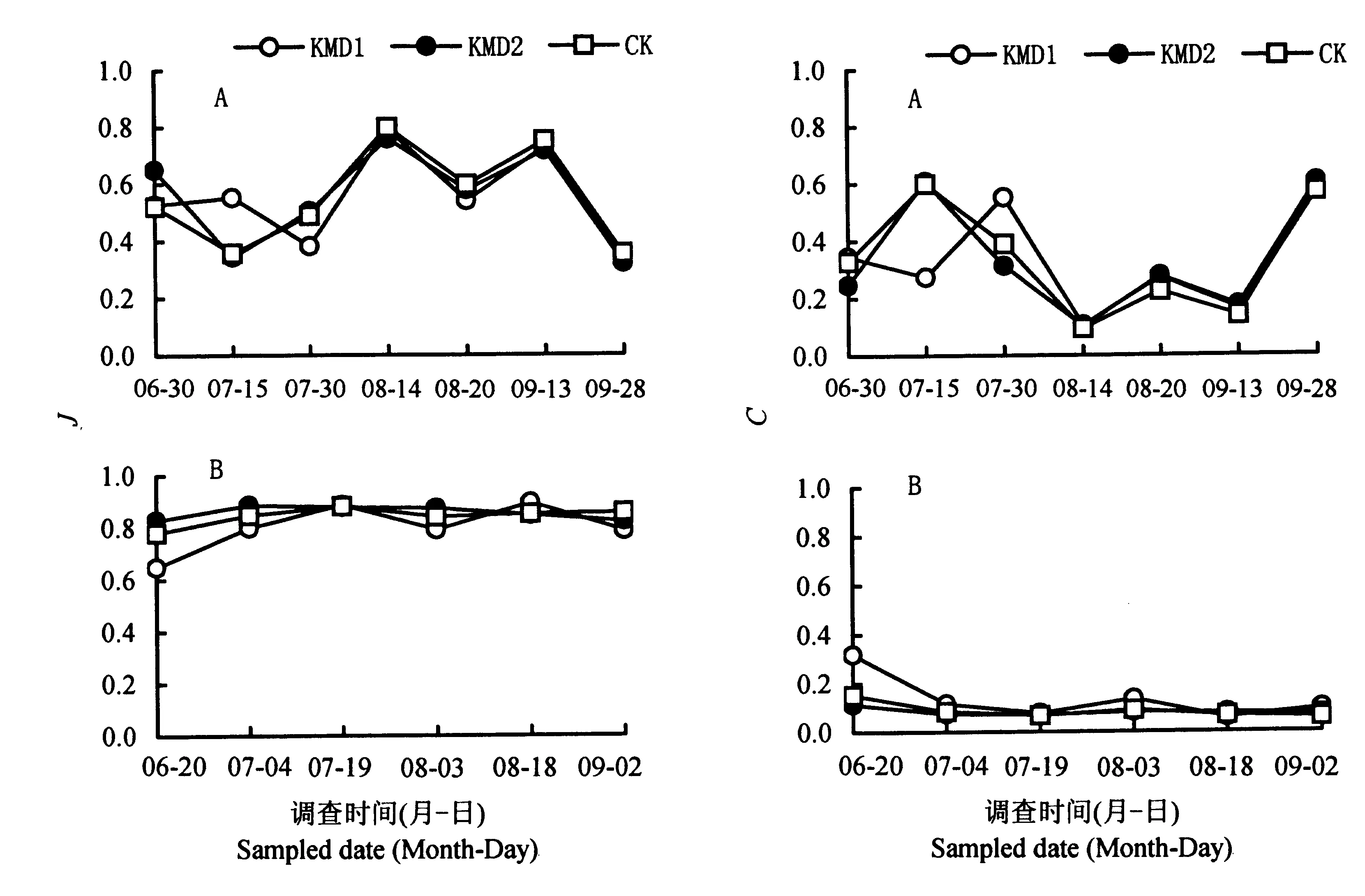
A: 杭州ⅠHangzhouⅠ; B: 杭州ⅡHangzhouⅡ。
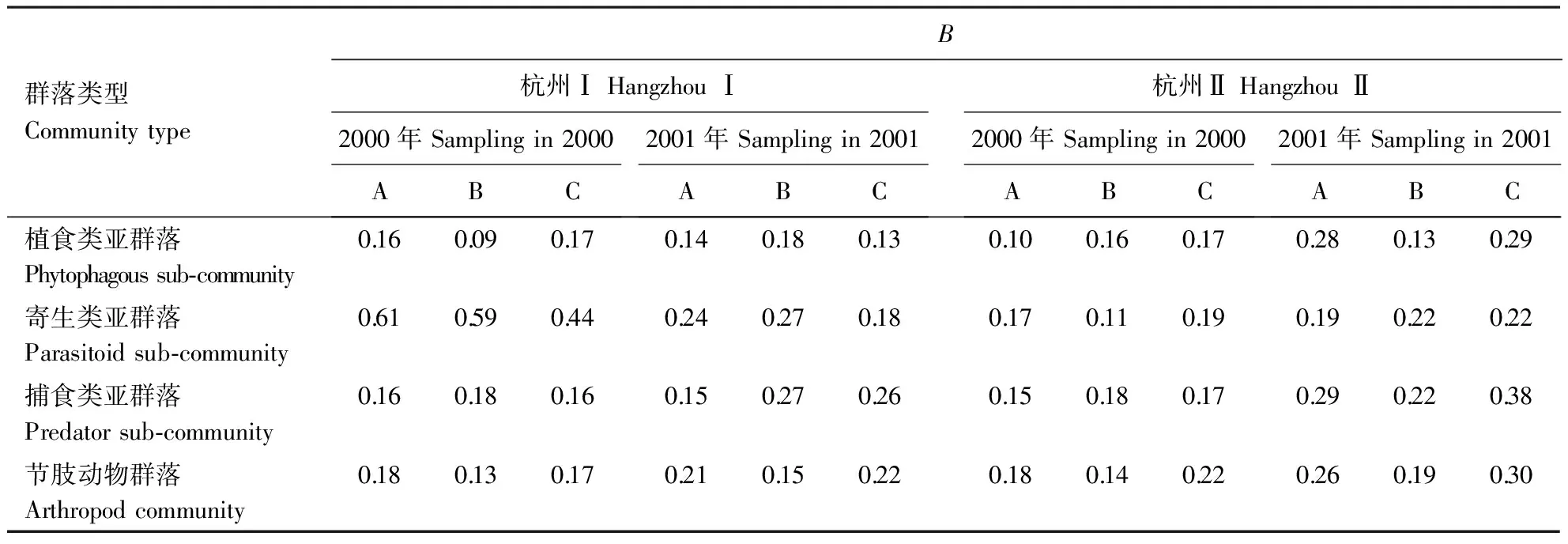
表3 转cry1Ab基因粳稻与对照间节肢动物群落的Bray-Curtis距离系数(B)
A、B分别表示KMD1、KMD2的亚群落或群落与对照间的比较,C表示KMD1与KMD2间亚群落或群落的比较。
A, B indicate comparison between the sub-communities or communities of KMD1, KMD2 and those of the control, respectively. C indicates comparison between the sub-communities or communities of KMD1 and those of KMD2.
KMD1与KMD2节肢动物群落结构间的Bray-Curtis距离系数介于Bt粳稻与对照间距离系数的变幅范围内或略高(表3)。这种差异可能与节肢动物在田块中的分布不完全均一而导致的取样误差有关。这也从一个侧面说明了Bt粳稻与对照间亚群落的相异性较低,其相异性可能在一定程度上是由取样误差所致。
3 讨论
本研究结果表明,Bt粳稻与对照间的各功能团优势度、群落结构参数(S、H′、J、C)及其时间动态在大多数情况下无明显差异;Bt粳稻与对照间植食类、寄生类、捕食类亚群落和整个节肢动物群落的相似性一般较高。故可认为,多数情况下Bt粳稻对稻田节肢动物群落结构无明显的负面影响。这可能与稻田中Bt粳稻的靶标害虫如螟蛾科和夜蛾科昆虫在植食类节肢动物功能团中的优势度较低,而非靶标害虫如飞虱科、叶蝉科的优势度较高有关。据我们2年3点调查结果表明,即使在对照区螟蛾科成虫优势度也仅为1.55%~3.87%,夜蛾科成虫优势度为0.31%~0.51%,而飞虱科和叶蝉科优势度各为12.24%~47.35%和17.72%~71.69%。正因如此,Bt粳稻对第二营养层靶标害虫及其天敌的作用在较大程度上被高优势度的第二营养层的非靶标害虫在节肢动物群落中的地位所弥补或掩盖,从而表现出对整个节肢动物无明显的负影响。这与Bt棉对昆虫群落、害虫和天敌亚群落有明显影响,导致亚群落或群落结构不如非转基因常规棉稳定的结果(崔金杰和夏敬源,2000)不同。这可能与Bt棉和Bt粳稻对其非靶标害虫发生影响的程度不一致有关。其中,Bt棉导致了棉蚜Aphisgossypii、白粉虱Trialeurodesvaporariorum、棉叶蝉Empoascabiguttula、棉盲蝽Lyguslucorum、棉蓟马Thripstabaci和朱砂叶螨Tetranychuscinnabrinus等非靶标害虫的发生明显加重(Luetal.,2010;崔金杰和夏敬源,1998、2000),进而使得这些害虫的天敌种类或数量发生变化(崔金杰和夏敬源,2000);而Bt粳稻(KMD1、KMD2)在多数情况下不会引起非靶标害虫飞虱和叶蝉种群数量的显著变化(Chenetal.,2007; 刘志诚等,2002;周霞等,2005),因此对其天敌种类或数量也不会产生明显的影响。
尽管总体而言,Bt粳稻对稻田节肢动物群落无明显的影响,但有时Bt粳稻稻田中寄生蜂的优势度低于对照(差异未达显著水平),亚群落结构与对照间的相异性也较高,这在一定程度上反映了Bt粳稻对寄生蜂可能会有负面影响。但这种负面作用不及Bt棉(崔金杰和夏敬源,2000)和Bt玉米(Obrycki,2001)明显,Bt棉和Bt玉米均能导致其对应的靶标害虫棉铃虫Helicoverpaarmigera和欧洲玉米螟Ostrinianubilalis的寄生蜂个体数显著减少。这可能与Bt粳稻靶标害虫(螟虫等)占整个节肢动物群落的比率较低有关。因此,有关Bt粳稻对各类寄生蜂的影响尚有待进一步深入研究,并宜对靶标害虫寄生蜂的优势种类从个体生长发育和种群动态角度做逐一评价分析。
此外,值得提出的是,本研究因生物安全管理的要求,试验小区面积相对较小,易出现小区间节肢动物的相互扩散,进而造成试验误差;同时,本研究只进行2年的试验,转基因水稻对生态环境的影响是一个长期的过程,故条件许可时,还有必要扩大试验面积和进行长期检测,以验证或完善本文的有关评价结果。
崔金杰,夏敬源.1998.麦套夏播转Bt基因棉田主要害虫及其天敌的发生规律.棉花学报,10(5):255-262.
崔金杰,夏敬源.2000.麦套夏播转Bt基因棉R93-6对昆虫群落的影响.昆虫学报,43(1):43-51.
郝树广,张孝羲,程遐年,罗跃进,田学志.1998.稻田节肢动物群落营养层及优势功能集团的组成与多样性动态.昆虫学报,41(4):343-353.
刘雨芳,张古忍,古德祥.1999.利用改装的吸虫器研究稻田节肢动物群落.植物保护,25(6):39-40.
刘志诚,叶恭银,胡萃,Swapan Kumar Datta.2002.Bt水稻对主要非靶标害虫和蜘蛛优势种田间种群动态的影响.植物保护学报,29(2):138-144.
唐启义,冯明光.1997.实用统计分析及其计算机处理平台.北京:中国农业出版社.
周霞,程家安,娄永根.2005.转cry1Ab基因水稻对非靶标昆虫白背飞虱种群增长的影响.昆虫学报,7(4):786-791.
Akhtar Z R, Tian J C, Chen Y, Fang Q, Hu C, Chen M, Peng Y F and Ye G Y. 2010. Impacts of sixBtrice lines on nontarget rice feeding thrips under laboratory and field conditions.EnvironmentalEntomology,39:715-726.
Bitzer R J, Rice M E, Pilcher C D, Plicher C L and Lam W K F. 2005. Biodiversity and community structure ofepedaphicandeuedaphicspringtails(Collembola) in transgenic rootwormBtcorn.EnvironmentalEntomology, 34:1346-1376.
Chen M, Liu Z C, Ye G Y, Shen Z C, Hu C, Peng Y F, Altosaar I and Shelton A M. 2007. Impacts of transgeniccry1Ab rice on non-target planthoppers and their main predatorCyrtorhinuslividipennis(Hemiptera: Miridae)—A case study of the compatibility ofBtrice with biological control.BiologicalControl, 42:242-250.
Chen Y, Tian J C, Shen Z C, Peng Y F, Hu C, Guo Y Y and Ye G Y. 2010. Transgenic rice plants expressing a fused protein of Cry1Ab/Vip3H has resistance to rice stem borers under laboratory and field conditions.JournalofEconomicEntomology, 103:1444-1453.
Datta K, Vasquez A, Tu J, Torrizo L, Alam M F, Oliva N, Abrigo E, Khush G S and Dara S K. 1998. Constitutive and tissue-specific differential expression of thecry1A(b) gene in transgenic rice plants conferring resistance to rice insect pest.TheoreticalandAppliedGenetics, 97(1-2):20-30.
Dhawan A K, Aneja A, Singh J and Saini S. 2009. Population dynamics of different pests onBt-cotton vis-a-vis meteorological parameters in Punjab.JournalofAgrometeorology, 11(2):180-182.
Head G, Moar M, Eubanks M, Freeman B, Ruberson J, Hagerty A and Turnipseed S. 2005. A multiyear, large-scale comparison of arthropod populations on commercially managedBtand non-Btcotton fields.EnvironmentalEntomology, 34:1257-1266.
Heong K L, Aquino G B and Barrion A T. 1991. Arthropod community structures of rice ecosystems in the Philippines.BulletinofEntomologicalResearch, 81(4):407-416.
James C. 2010. 2009 ISAAA report on global status of Biotech/GM crops.http://www.isaaa.org/.
Lu Y H, Wu K M, Jiang Y Y, Xia B, Li P, Feng H Q, Wyckhuys K A G and Guo Y Y. 2010. Mirid bug outbreaks in multiple crops correlated with fiide-scale adoption ofBtcotton in China.Science, 328:1151-1154.
Obrycki J J. 2001. Transgenic insecticidal corn: the agronomic and ecological rationale for its use-response.Bioscience, 51:903-905.
Riddick E W and Barbosa P. 1998. Impact of Cry3A-intoxicatedLeptinotarsadecemlineata(Coleoptera: Chrysomelidae) and pollen on consumption, development, and fecundity ofColeomegillamaculata(Coleoptera: Coccinellidae).AnnalsoftheEntomologicalSocietyofAmerica, 91:303-307.
Schoenly K G, Justo H D, Barrion A T, Harris M K and Bottrell D G. 1998. Analysis of invertebrate biodiversity in a Philippine farmer′s irrigated rice field.EnvironmentalEntomology, 27:1125-1136.
Shu Q Y, Ye G Y, Cui H R, Cheng X Y, Xiang Y B, Wu D X, Gao M W, Xia Y W, Hu C, Sardana R and Ailosaar I. 2000. Transgenic rice plants with a syntheticcry1Ab gene fromBacillusthuringiensiswere highly resistant to eight lepidopteran rice pest species.MolecularBreeding, 6:433-439.
Sisterson M S, Biggs R W, Olson C, Carriere Y, Dennehy T J and Tabashnik B E. 2004. Arthropod abundance and diversity inBtand non-Btcotton fields.EnvironmentalEntomology, 33:921-929.
Tian J C, Liu Z C, Chen M, Chen Y, Chen X X, Peng Y F, Hu C and Ye G Y. 2010. Laboratory and field assessments of prey-mediated effects of transgenicBtrice onUmmeliatainsecticeps(Araneida: Linyphiidae).EnvironmentalEntomology, 39:1369-1377.
Tu J M, Zhang G A, Datta K, Xu C G, He Y Q, Zhang Q F, Khush G S and Datta S K. 2000. Field performance of transgenic elite commercial hybrid rice expressingBacillusthuringiensisdelta-endotoxin.NatureBiotechnology, 18:1101-1104.
Whitehouse M E A, Wilson J and Fitt G P. 2005. A comparison of arthropod communities in transgenicBtand conventional cotton in Australia.EnvironmentalEntomology, 34:1224-1241.
Ye G Y, Shu Q Y, Yao H W, Cui H R, Cheng X Y, Hu C, Xia Y W, Gao M W and Altosaar I. 2001. Field evaluation of resistance of transgenic rice containing a syntheticcry1Ab gene fromBacillusthuringiensisBerliner to two stem borers.JournalofEconomicEntomology, 94:271-276.
Ye G Y, Yao H W, Shu Q Y, Cheng X, Hu C, Xia Y W, Gao M W and Altosaar I. 2003. High levels of stable resistance in transgenic rice with acry1Ab gene fromBacillusthuringiensisBerliner to rice leaffolder,Cnaphalocrocismedinalis(Guenée) under field conditions.CropProtection, 22:171-178.
Impactoftransgeniccry1AbjaponicariceonthearthropodcommunityofricepaddiesinChina
Zhi-cheng LIU1, Yang CHEN1, Jun-ce TIAN1, Zeng-bin LU1, Qing-yao SHU2, Cui HU1, Yu-fa PENG3, Gong-yin YE11
InstituteofInsectSciences,StateKeyLaboratoryofRiceBiology,MinistryofAgricultureKeyLaboratoryofMolecularBiologyofCropPathogensandInsects,ZhejiangUniversity;2InstituteofNuclearAgriculturalSciences,ZhejiangUniversity,Hangzhou,Zhejiang310029,China;3StateKeyLaboratoryforBiologyofPlantDiseasesandInsectPests,InstituteofPlantProtection,ChineseAcademyofAgriculturalSciences,Beijing100193,China
The community of arthropods in rice paddies was sampled at three locations over two years. The arthropods captured were divided into five feeding guilds: phytophages, parasitoids, predators, detritivores and others. The effects of two transgenicBacillusthuringiensiscry1Abjaponicalines (KMD1 and KMD2) were evaluated in terms of guild dominance, family composition and dominance, as well as several indicdes of diversity. The control community was the one present on the untransformed parental rice line Xiushui 11. Compared with the control paddies, the guild dominance, species richness (S), Shannon-Wiener′s diversity index (H′) , evenness index (J) and Simpson′s dominance index (C), and the temporal dynamics ofBtrice paddies showed no significant difference at the most situation. The dissimilarities between phytophagous sub-communities, parasitoid sub-communities, predator sub-communities and arthropod communities as a whole inBtrice plots and the control were apparently low. It was apparent that the transgeniccry1Abjaponicarice has no negative effect on the rice paddy arthropod community.
Btjaponicarice;cry1Ab gene; arthropod; guild; community
2011-01-16接受日期2011-02-10
国家重大基础研究发展计划(973)(2007CB109202);国家转基因生物培育重大专项(2008ZX08011-01)
叶恭银,E-mail:chu@zju.edu.cn
