人骨肉瘤细胞MG-63抗失巢凋亡的机制研究*
2011-10-24林丁盛张小磊蒋良福高伟阳
林丁盛, 张小磊, 蒋良福, 高伟阳
(温州医学院附属第二医院骨科, 浙江 温州 325000)
人骨肉瘤细胞MG-63抗失巢凋亡的机制研究*
林丁盛, 张小磊, 蒋良福, 高伟阳△
(温州医学院附属第二医院骨科, 浙江 温州 325000)
目的观察人骨肉瘤细胞(MG-63)是否存在抗失巢凋亡(anoikis)的特性,并进一步研究其分子机制。方法将MG-63细胞分别在普通6孔板和事先用聚甲基丙烯酸2-羧乙基酯(poly-HEMA)处理过的6孔板中培育24 h、48 h、72 h和7 d,人正常成骨细胞hFOB 1.19和人正常肾上皮细胞293T作对照,在光镜、电镜下观察细胞的形态以及细胞间连接,用流式细胞术检测细胞的凋亡率。通过RT-PCR方法检测不同时期、不同生长状态下,细胞中β-catenin和磷脂酰肌醇激酶(PI3K)的转录水平,并用Western blotting方法检测细胞内β-catenin、PI3K、Bcl-2和caspase-3、8、9的蛋白表达。结果MG-63细胞在悬浮状态下培养,细胞相互之间聚集成比较致密的团块,没有发生明显的细胞凋亡(凋亡比例最高为30.29%)。在贴壁生长和悬浮生长情况下,caspase-3和caspase-9的活化水平均无显著差异,而在悬浮状态下caspase-8在48 h时活化最多,然后活性减低,β-catenin和磷脂酰肌醇激酶的转录水平比对照组高,但蛋白表达水平对照组与实验组无显著差异。在悬浮状态下,Bcl-2的蛋白表达水平呈时间依赖性增长,在72 h达到最高。结论MG-63细胞具有抗失巢凋亡的特性。细胞从细胞外基质中脱离后,早期可能激活caspase-8,从而启动并执行anoikis;Bcl-2的活化可能在后期抑制anoikis的发生。
失巢凋亡; 骨肉瘤 ; 蛋白质Bcl-2; 半胱氨酸天冬氨酸蛋白酶8
正常的上皮细胞是黏附依赖的,它们的存活需要来自基质的信号,如果失去基质的支持,细胞会发生程序性死亡,称为失巢凋亡(anoikis)[1-3]。失巢凋亡的意义在于防止这些脱落的细胞种植于其它不适当的地方继续生长,然而很多容易发生转移的恶性肿瘤细胞,从瘤体上脱落后并不发生凋亡,而且可以迁移到其它部位再次生长,这种抗失巢凋亡的现象,被认为是肿瘤发生转移的重要原因之一。骨肉瘤是常见的恶性骨肿瘤,本文旨在研究骨肉瘤细胞是否存在抗失巢凋亡的特性,并进一步研究其分子机制,以期对骨肉瘤细胞的治疗与预防提供新的治疗思路。
材 料 和 方 法
1主要试剂及仪器
聚甲基丙烯酸2-羧乙基酯(poly-2-hydroxyethyl methacrylate,poly-HEMA)、兔抗人caspase-3单克隆抗体购自Sigma。鼠抗人β-catenin、磷脂酰肌醇3-激酶(phosphoinositide3-kinase,PI3K)85、PI3K 110、Bcl-2和caspase-9单克隆抗体均购自Santa Cruz。鼠抗人caspase-8单克隆抗体购自Calbiochem。山羊抗小鼠IgG-HRP和山羊抗兔IgG-HRP为北京中山生物技术公司产品。逆转录聚合酶链反应试剂购自上海生物工程公司。细胞培养瓶和细胞培养液购自杭州四季青生物工程公司。FAC Scan 流式细胞仪为Becton Dickinson产品。垂直板电泳槽电转移仪由Bio-Rad生产。电子显微镜为 Philps Tecnal 10型。
2细胞系
人骨肉瘤细胞MG63从武汉典藏中心购得,对照细胞系人成骨细胞hFOB 1.19,人肾上皮细胞293T细胞购自上海中科院细胞研究所,MG63细胞与293T细胞于DMEM培养液,hFOB 1.19细胞于1∶1 F12/DMEM+McCoy’s 5A(Invitrogen GibcoTM)培养液中培养,加上10%灭活的胎牛血清、2 mmol/L L-谷氨酰胺、1×105U/L青霉素和1×105U/L链霉素,在5% CO2、37 ℃培养箱中培养。
3细胞悬浮培养皿的准备
将2 mL 50 g/L的poly-HEME无水乙醇溶液加入6孔板中,置室温待其完全干燥,重复上述步骤1次,然后使用前用1×PBS缓冲液将6孔细胞培养板冲洗4遍,置于超净台中紫外线照射消毒待用。
4细胞的处理
将细胞用含0.01% EDTA的0.25%胰蛋白酶进行消化,制成约5×108cells/L的单细胞悬液,将2 mL接种于poly-HEMA处理过的6孔板中,同时取2 mL接种于普通的6孔板中作为阴性对照,将细胞置于37 ℃、5% CO2培养箱中培养24 h、48 h、72 h及7 d。
5光学显微镜下观测细胞形态
取实验组和对照组细胞各5×105细胞,加入2 mL培养液置于光学显微镜下观察细胞形态,以及细胞集落形成情况。
6电子显微镜观测细胞形态及细胞结构
按常规电镜样品制备技术操作。固定,收集细胞,用体积分数为2%戊二醛固定过夜;后固定,离心弃固定液,PBS洗涤2次后,用质量浓度为1%锇酸后固定1 h;预染,离心弃固定液,蒸馏水洗涤2次后,用质量浓度为2%醋酸铅预染1 h;脱水,离心弃液预染,蒸馏水洗涤2次后,依次用体积分数35%、50%、75%、90%乙醇梯度脱水;环氧树脂(Epon812)包埋;LKB超薄切片机切片;柠檬酸铅染色;透色电镜观察并摄片(80 kV)。
7流式细胞仪检测细胞凋亡率
取5×108cells/L细胞,70%冰乙醇固定,于4 ℃冰箱保存24 h,24 h 后PBS 洗2次,加入1 g/L RNase A 200 μL,37 ℃水浴30 min,再加PI 染色液避光反应30 min,上机检测,将所得数据采用专用软件CellQuest计算细胞凋亡比例。
8RT-PCR
5×108cells/L细胞加入1 mL Trizol 试剂,按照说明书操作提取总RNA,测定260和280 nm 处的吸光度,估计纯度并定量RNA。RT:反应体积为20 μL;组成为:dNTPs 2 μL,5×逆转录buffer 4 μL,RNasin 20 U,Mo-AMV逆转录酶200 U,随机六聚引物2 μg,总RNA 2 μg,去离子水加至20 μL。25 ℃ 10 min,42 ℃ 1 h,70 ℃ 10 min,停止反应。 PCR:反应体积为50 μL;组成为:10×PCR buffer 5 μL,Taq DNA酶2.5 U,逆转录产物2 μL,MgCl22 mmol/L,β-actin 正负链引物各0.2 μg,β-catenin和PI3K正负链引物各0.4 μg,去离子水加至50 μL。94 ℃预变性5 min,再94 ℃ 30 s,适当退火温度1 min,72℃ 1 min,30个循环,72 ℃ 10 min 结束。用2%琼脂糖凝胶电泳分离反应产物,溴化乙啶染色显示,通过紫外成像系统对凝胶中各产物条带吸光度进行积分,用细胞因子与β-actin PCR产物的吸光度绝对量比值对细胞因子的表达量进行相对定量。引物序列: 人β-catenin,正义引物 5’-GATTTGATGGAGTTGGACATGG-3’,反义引物 5’-TGTTCTTGAGTGAAGGACTGAG-3’;人PI3K,正义引物 5’-ATTGTGATACACCCTCCGTGGA-3’,反义引物 5’-TGATGAGGTATGCTAGGCGACC-3’;人GAPDH,正义引物 5’-ACCTGACCTGCCGTCTAGAA-3’,反义引物 5’-TCCACCACCCTGTTGCTGTA-3’。
9蛋白免疫印迹(Westernblotting)分析相关蛋白的变化
收集细胞,溶于1 mL RIPA细胞裂解液中[50×10-3mol/L Tris-HCl(pH 7.4),150×10-3mol/L NaCl,1% NP40,5×10-3mol/L EDTA,5×10-3mol/L NaF,2×10-3mol/L Na3VO4,1×10-3mol/L PMSF,5 mg/L leupeptine,5 mg/L aprotinin]。Bradford法检测蛋白浓度,将各组浓度调到同一水平,经12%SDS-PAGE电泳,将分离的蛋白转移至PVDF膜,用5%脱脂奶粉封闭过夜,加入β-catenin、Bcl-2、PI3K 85 、PI3K 110、caspase-3、8、9单抗和β-actin(1∶1 000),摇床上振摇2 h,PBST洗涤3次,每次10 min。再加入抗小鼠IgG-HRP,摇床上振摇2 h,PBST洗涤3次,ECL试剂显色,干燥,显影在柯达X光片上。在Leica Q500iw 图像分析软件上半定量分析条带灰度值。
10统计学处理
结 果
1光镜下细胞贴壁培养与悬浮培养的形态学表现
hFOB 1.19、293T、MG-63细胞贴壁状态下生长良好,但hFOB 1.19、293T细胞在悬浮培养下细胞之间松散,几乎不形成细胞聚集体,然而MG-63细胞相互之间聚集成比较大而紧密的聚集体,散在的细胞很少,见图1。
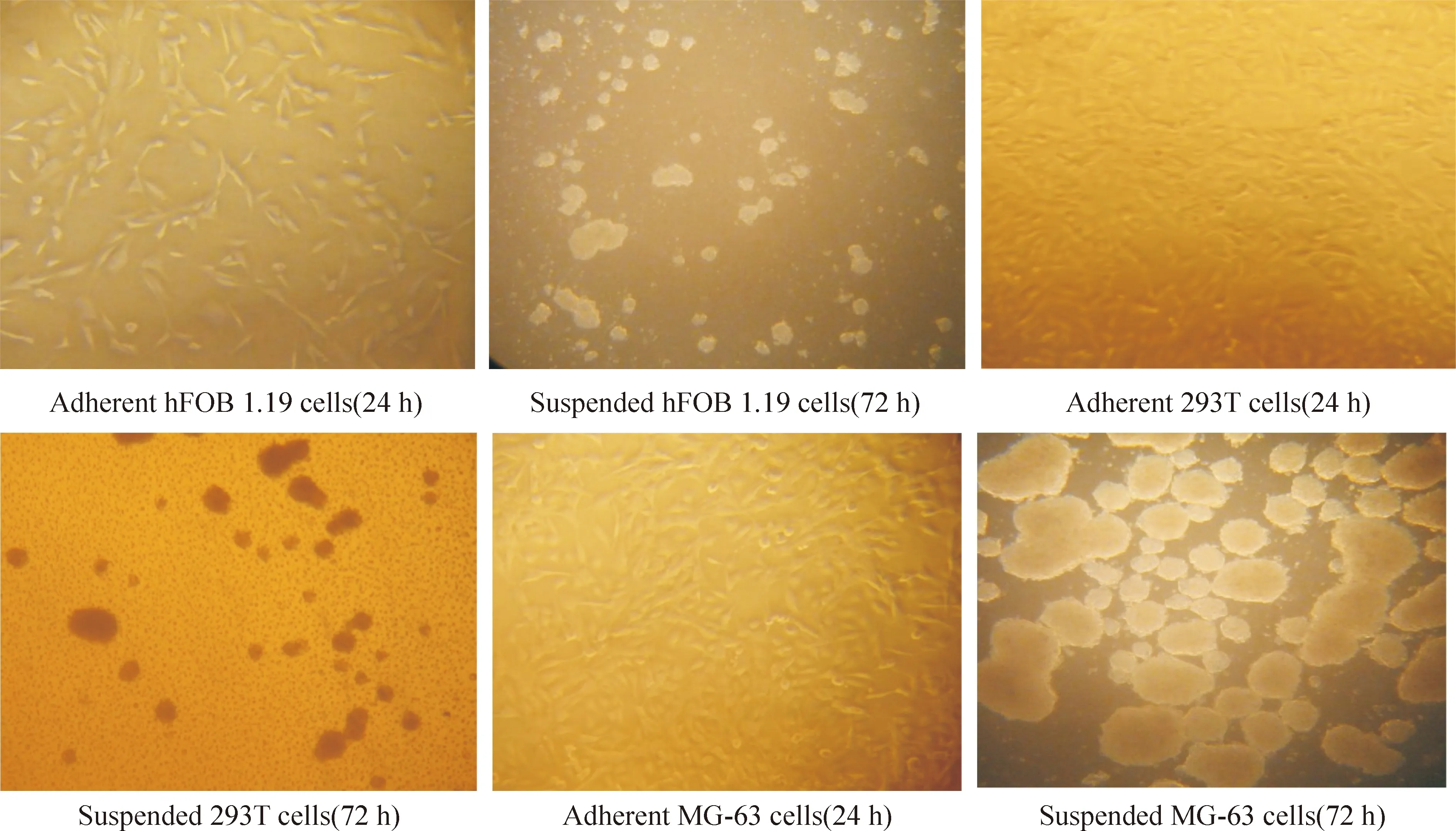
Figure 1. Cell morphology of MG-63, 293T and hFOB 1.19 cells in poly-HEMA treated (in suspension condition) wells (×40 ) and poly-HEMA untreated (in adherent condition) wells (×100) under light microscope. The background of the suspended cell culture wells is poly-HEMA.
图1光镜下细胞贴壁培养与悬浮培养的形态学表现
2电镜下细胞悬浮培养的形态学及细胞结构观察
细胞悬浮培养72 h,电镜下发现hFOB 1.19细胞以及293T细胞发生凋亡,形态学上表现凋亡的特性,但大部分MG-63细胞正常生长,细胞保持正常状态。所有的细胞都建立起缝隙连接和桥粒等细胞间的连接,见图2。
3细胞贴壁培养与悬浮培养的凋亡率
在悬浮状态下培养,MG-63细胞的凋亡率明显低于hFOB 1.19细胞和293T细胞的凋亡率。MG-63细胞在贴壁生长24 h后,其凋亡率为(3.17±0.75)%,悬浮状态下培养24 h、48 h、72 h和7 d后,凋亡率分别为(6.27±2.88)%、(13.91±1.50)%、(20.33±1.62)%、(30.29±2.60)%,而hFOB 1.19细胞和293T细胞在悬浮状态下生长7 d的凋亡率分别是(89.23±3.71)%、(95.88±1.32)%。 MG-63细胞具有抵抗失巢凋亡的能力,见图3。
Figure 2. Cell morphology and cell junctions of MG-63, 293T and hFOB 1.19 cells in poly-HEMA treated wells under transmission electronic microscope.Cells were seeded on poly-HEMA untreated (in adherent condition) wells for 24 h and poly-HEMA treated (in suspension condition) wells for 72 h and were collected for TEM observation. Cells morphology were photographed(×3 700). Bar:5 μm. Cell junctions (suspended 72 h) were also photographed(×65 000). Bar:0.2 μm.
图2电镜下细胞悬浮培养的形态学及细胞结构观察

图3不同时间细胞贴壁生长与悬浮生长的凋亡率
4贴壁培养与悬浮培养对MG63骨肉瘤细胞PI3K和β-cateninmRNA和蛋白表达的影响
在悬浮培养24 h后,β-catenin 基因转录水平较贴壁状态时减弱,但随后β-catenin基因转录呈时间依赖性的增长,但β-catenin的蛋白表达在贴壁状态和悬浮状态下无显著差异。在悬浮状态下PI3K在24 h和48 h的转录水平比对照组高。PI3K蛋白有2个亚基,分别为P85 和 P110。其中无功能性的PI3K 85蛋白表达在72 h达到最高。PI3K 110蛋白表达对照组与实验组无显著差异,见图4。
5贴壁培养与悬浮培养对MG63骨肉瘤细胞Bcl-2和caspase-3、8、9蛋白表达的影响
在悬浮状态下培养24 h、48 h和72 h后,caspase-3和caspase-9蛋白无活化,与贴壁状态下培养24 h无显著差异。而在悬浮状态下caspase-8在48 h时活化最多,Bcl-2的蛋白表达水平呈时间依赖性增长,在72 h达到最高,见图5。

图4贴壁培养条件与悬浮培养条件下MG63细胞PI3K和β-cateninmRNA和蛋白的表达

Figure 5. The translational level of caspase-3, caspase-8, caspase-9 and Bcl-2 in MG-63 cells seeded on poly-HEMA treated wells.1: MG-63 cell cultured in poly-HEMA untreated wells for 24 h. 2, 3, 4: MG-63 cell cultured in poly-HEMA treated wells for 24 h, 48 h and 72 h,respectively.n=3.
图5贴壁培养与悬浮培养条件下MG63骨肉瘤细胞Bcl-2和caspase-3、8、9蛋白表达情况
讨 论
很多类型细胞必须与胞外基质(ECM)黏附才能增殖,这种现象称为锚定依赖,这些细胞一旦与基质分离,使外界输入的存活信息中断就引发失巢凋亡。本实验发现,hFOB 1.19和293T细胞在细胞与细胞基质分离后,立即发生失巢凋亡,而MG-63细胞在悬浮状态下培养7 d,最高的凋亡率为(30.29±2.60)%,表现出较强的抗凋亡特性,从而为骨肉瘤的远处转移和局部侵袭创造条件。
细胞同源性的黏附和细胞之间的信号转导,对细胞的生物学性状、生长、凋亡和分化有着重要的作用,同时对肿瘤细胞的侵袭性和转移也有着重要的作用[4-7]。我们发现不同的细胞在悬浮状态下形成不同的聚集体,MG-63细胞在悬浮状态下相互之间形成了大而致密的聚集体,没有发生明显的细胞凋亡,而hFOB 1.19和293T细胞在悬浮培养下细胞之间松散,几乎不形成细胞聚集体,发生明显的anoikis,这可能MG-63细胞虽然失去与ECM的接触,但仍会有细胞与细胞之间的连接,从而通过细胞与细胞之间信号转导,模拟细胞与基质之间的信号转导,从而使悬浮的细胞逃避anoikis[7-9]。
细胞与细胞之间的连接包括缝隙连接、桥粒等。缝隙连接存在于大部分组织和细胞间,桥粒通常只存在于上皮细胞,但我们电镜观察,hFOB 1.19和293T细胞和MG-63细胞在悬浮状态下都形成了桥粒结构,这可能悬浮状态下培养刺激细胞与细胞间连接的形成,但对细胞存活或凋亡的调控还依赖于不同存活或者凋亡信号的转导。
β-catenin是细胞与细胞锚联的重要信号分子,它可以通过信号转导细胞增殖和转化,激活PI3K,通过PI3K/Akt磷酸化而转导细胞存活信号,从而抑制凋亡[7,10,11],但我们发现虽然在悬浮状态下β-catenin和PI3K的转录水平比对照组高,但蛋白表达水平对照组与实验组无显著差异,这可能存在某些分子抑制了β-catenin和PI3K的蛋白翻译或者降解了β-catenin和PI3K蛋白。
Caspase是执行细胞凋亡的主要酶类,活化的caspase通过特异性的裂解底物而导致细胞的凋亡,通常caspase-8、9被认为是凋亡起始因子,caspase-3作为凋亡的执行因子[12]。吕海等[13]报道,人干扰素α2a可以通过调节细胞caspase信号途径,诱导骨肉瘤细胞MG-63产生anoikis现象。有文献报道[7,11,14,15],当细胞与基质分离时,可以激活caspase-8,同时过度的表达Bcl-2可以抑制caspase-8的活化。我们实验中,在悬浮状态下,Bcl-2蛋白呈时间依赖性增长表达,caspase-8在起始48h就被活化,然后活性降低,但caspase-3活化水平始终无显著改变,这可能在细胞悬浮早期细胞与基质分离或者其它一些通路激活了caspase-8,但高水平的Bcl-2表达以及可能一些其它分子表达,抑制了caspase-8的促凋亡能力,从而使促凋亡和抑凋亡达到了新的平衡,使MG63骨肉瘤细胞逃避anoikis。
[1] Liotta LA, Kohn E. Anoikis: cancer and the homeless cell[J]. Nature,2004,430(7003):973-974.
[2] Frisch SM,Francis H. Disruption of epithelial cell-matrix interactions induces apoptosis[J]. J Cell Biol, 1994, 124(4):619-626.
[3] Bretland AJ, Lawry J,Sharrard RM. A study of death by anoikis in cultured epithelial cells[J]. Cell Prolif,2001,34(4):199-210.
[4] Steuer AF, Rhim JS, Hentosh PM, et al. Survival of human cells in the aggregate form: potential index ofinvitrocell transformation[J]. J Natl Cancer Inst, 1977, 58(4):917-921.
[5] Frisch SM,Ruoslahti E. Integrins and anoikis[J]. Curr Opin Cell Biol,1997, 9(5):701-706.
[6] Frisch SM, Screaton RA. Anoikis mechanisms[J]. Curr Opin Cell Biol, 2001,13(5):555-562.
[7] Grossmann J. Molecular mechanisms of “detachment-induced apoptosis-Anoikis”[J]. Apoptosis, 2002,7(3):247-260.
[8] Zhang Y, Lu H, Dazin P,et al Squamous cell carcinoma cell aggregates escape suspension-induced, p53-mediated anoikis: fibronectin and integrin αvmediate survival signals through focal adhesion kinase[J]. J Biol Chem, 2004, 279(46):48342-48349.
[9] Shen X,Kramer RH. Adhesion-mediated squamous cell carcinoma survival through ligand-independent activation of epidermal growth factor receptor[J]. Am J Pathol, 2004, 165(4):1315-1329.
[10]Fuchs SY,Ougolkov AV,Spiegelman VS,et al. Oncogenic β-catenin signaling networks in colorectal cancer[J]. Cell Cycle, 2005,4(11):1522-1539.
[11]Reddig PJ, Juliano RL. Clinging to life: cell to matrix adhesion and cell survival[J]. Cancer Metastasis Rev, 2005, 24(3):425-439.
[12]方 勇,吴金民,潘宏铭.Caspase-3抑制剂抑制长春碱诱导的人乳腺癌细胞凋亡及IκB-α降解[J].中国病理生理杂志,2009,25(2):215-219.
[13]吕 海, 郑燕芳, 金大地. 人α干扰素诱导骨肉瘤细胞Anoikis凋亡[J]. 中国肿瘤生物治疗杂志,2003 ,10(1):34-38.
[14]Frisch SM. Evidence for a function of death-receptor-related, death-domain-containing proteins in anoikis[J]. Curr Biol, 1999,9(18):1047-1049.
[15]Park MY, Lee RH, Lee SH, et al. Apoptosis induced by inhibition of contact with extracellular matrix in mouse collecting duct cells[J].Nephron, 1999,83(4):341-351.
PossiblemechanismofanoikisresistanceinhumanosteosarcomaMG-63cells
LIN Ding-sheng, ZHANG Xiao-lei, JIANG Liang-fu, GAO Wei-yang
(DepartmentofOrthopaedicSurgery,SecondAffiliatedHospitalofWenzhouMedicalCollege,Wenzhou325000,China.E-mail:linden2008@yahoo.com.cn)
AIM: To investigate the possible mechanism for the regulation of anchorage-independent survival by observing the resistance of human osteosarcoma cell line MG-63 to anoikis.METHODSHuman osteosarcoma cell line MG-63 was cultured in six-well plates under the conditions of pretreating with or without poly-2-hydroxyethyl methacrylate(poly-HEMA) for 24 h, 48 h, 72 h and 7 days. Human fetal osteoblasts cell hFOB 1.19 and human kidney epithelial cell 293T were used as controls. Morphological changes of the cells treated with ploy-HEMA were observed under microscope by aggregation assay. The cell junctions were evaluated under transmission electronic microscope. Apoptosis rate in response to anoikis was determined by flow cytometry. The transcriptional levels of β-catenin and phosphoinositide 3-kinase(PI3K) were analyzed by RT-PCR. The protein levels of β-catenin, PI3K, Bcl-2 and activation of caspase-3, 8, 9 were examined by Western blotting.RESULTSHuman kidney epithelial cell 293T and human osteoblast cell hFOB1.19 significantly underwent anoikis when adherence was denied. Human osteosarcoma MG-63 cells were distinctly anoikis-resistant when detached. Caspase-8 in suspension cultured MG-63 cells was activated by cell-matrix detachment. Translational level of Bcl-2 significantly increased in a time-dependent manner.CONCLUSIONThe MG-63 cells show resistance to anoikis when detachment of adherent cells from extracellular matrix occurs. Over-expression of Bcl-2 participates in the process of anoikis by blocking the activation of caspase-8, resulting in the resistance of MG-63 cells to anoikis.
Anoikis; Osteosarcoma; Protein Bcl-2; Caspase-8
R73-3
A
10.3969/j.issn.1000-4718.2011.01.032
1000-4718(2011)01-0165-06
2010-07-08
2010-11-08
浙江省自然科学基金资助项目(No. Y2100897);温州市科技计划资助项目(No.Y20100233)
△ 通讯作者Tel 0577-88879012; E-mail: linden2008@yahoo.com.cn
