福寿螺的寄主植物及其对福寿螺体重的影响
2011-10-12叶建人冯永斌林贤文王华弟祝增荣
叶建人,冯永斌,林贤文,,王华弟,祝增荣
1浙江省温岭市植保站,浙江温岭317500;2浙江大学昆虫科学研究所,水稻生物学国家重点实验室,浙江杭州310029;3浙江省植物保护检疫局,浙江杭州310020
福寿螺Pomacea canaliculata Lamarck,又称大瓶螺、苹果螺、雪螺,隶属瓶螺科Ampullariidae瓶螺属Pomacea,原产于南美洲亚马逊河流域,是世界100种恶性外来入侵物种之一(Naylor,1996;Joshi&Sebastian,2006),也是我国公布的第一批(16种)外来入侵物种(环保总局和中国科学院,2003)之一。其适应性强、食性杂、繁殖率高、生长迅速,繁殖速度比亚洲稻田中本地近缘物种快10倍左右,自入侵以来很快在南方大部分地区形成自然种群,严重威胁入侵地水域及附近的生物多样性和生态系统,危害农作物(水稻、茭白、白莲等)及其他水生植物(菱角等),造成巨大的经济损失(尹绍武等,2000;俞晓平等,2001;陈建明等,2003;叶建人和李云明,2009;叶建人等,2011)。由于其寄主范围广,可以快速扩散蔓延;在浙江沿海,福寿螺以每年8~10 km的速度从南向北自然扩展(俞晓平等,2001),已在众多地区自然入侵后暴发为害(杨叶欣等,2010)。
研究福寿螺对广大水域中常见植物的寄主选择性,确定植物与福寿螺之间的营养联系,分析其取食与生长之间的关系及影响因子,将有助于我们了解其入侵的过程及扩散传播规律(Morrison&Hay,2011),是生物入侵研究中要解决的核心问题之一(万方浩等,2009),可以为建立监测预警技术和方法、制定合理的防治措施提供参考。同时可以为其生态控制和综合防治提供重要的生态学依据和技术支撑(章家恩和方丽,2008)。因此,本研究对浙江省温岭市水稻苗、菖蒲、异型莎草、菹草和苦瓜等典型的稻田水域及周边随处可见的24科43种植物进行调查,以了解福寿螺的寄主选择性及其取食后的发育情况。
1 材料与方法
1.1 试验材料
试验时间为2009年5~8月,在浙江省农作物病虫监测网温岭区域站病虫观测场(温岭市横峰街道后洋郑村)内进行。螺源为场内专设的福寿螺饲养池中的越冬代和1代成螺(>6.5 g·只-1);供试植物(样品)于温岭市郊水稻田、茭白田、池塘、沟渠等直接采集,部分从市场上购买,除辣椒、苦瓜用果实及藕用块茎外,原则上选用处于营养生长盛期并未进入开花生殖期的正常(健壮)植株用于试验。在场内挖掘长20 m、宽50 cm、深40 cm的水沟,沟内放置长35 cm、宽35 cm、高60 cm的饲养塑料网(25目)箱,灌20~30 cm水层,保证网箱内的福寿螺无法逃逸,也无外源福寿螺进入。水沟内的水通过灌排水流动,每天更换并清除排泄物。
1.2 试验方法
本试验属于非选择性试验(no-choice assay)。当天采集到的受试植物(样品)除辣椒、苦瓜外,其他需在去掉枯死的茎叶后,以相同数量的整株数为原则喂食,个别较易被取食的样品适当加量。将样品分成6份,称重后3份放入处理饲养网箱,3份放入对照的饲养网箱中,然后在饲养池内取30只成螺,分成3份,每份10只,分别称重后随机放入其中3只网箱内让其自然啮食。该步骤在0.5 h内完成,以保证成螺的饥饿程度基本一致,另外3个作为各自样品处理的对照。每天观察啮食情况,隔天将饲养网箱放入另外干净的水沟并进行适当的清理。分别在放置3、5 d时对受试植物(样品)进行称重,以计算试样相对减少率;放置5 d时称成螺重量,计算每日每只成螺体重增长率。
1.3 数据分析
受试植物(样品)减少率/%=[放螺前受试植物(样品)重量-放螺后受试植物(样品)重量]/放螺前受试植物(样品)重量×100
受试植物(样品)相对减少率/%=[放螺区受试植物(样品)减少率-不放螺区受试植物(样品)减少率]/[1-不放螺区受试植物(样品)减少率]×100
每日每只成螺体重增长率/%=[啮食受试植物(样品)后成螺体重-啮食受试植物(样品)前成螺体重]/啮食受试植物(样品)前成螺体重×100
在DPS 9.50版数据处理系统(唐启义,2010)中,采用LSD法多重比较处理间的差异;用直线回归法建立反正弦平方根转换后的相对取食率与成螺增长率之间的关系。根据相关文献测定的多种植物的总酚含量,与本试验测定的福寿螺的生长率做相关分析。
2 结果与分析
2.1 福寿螺的寄主范围
结果表明,福寿螺均能啮食试验中选用的24科43种植物(样品),其中包括水稻、茭白、莲藕及慈姑等15种常见的农作物,以及稗草、千金子、鸭舌草、矮慈菇等28种常见杂草。具体如下:
(1)槐叶苹科Salviniaceae:槐叶萍Salvinia nutans(L.)All.。(2)眼子菜科 Potamogetonaceae:菹草Potamogeton crispus L.。(3)禾本科 Poaceae:水稻苗 Oryza sativa L.,茭白苗 Zizania caduciflora(Turcz.et Trin.)Hand,马唐 Digitaria sanguinalis(L.)Scop,稗草 Echinochloa crusgalli(L.)Beauv,棒头草 Polypogon fugax Nees ex Steud,罔草 Beckmannia syzigachne(Steud.)Fern,千金子 Semen Euphorbiae Lathyridis,双穗雀稗 Paspalum paspaloides(Michx.)Scribn,芦苇 Phragmites australis(Cav.)Trin.ex Steud,纤毛鹅观草 Roegneria ciliaris(Trin.)Nevski,黑麦草 Lolium perenne L.,早熟禾 Poa annua L.,牛筋草 Eleusine indica(L.)Gaerth,水稗 Echinochloa phyllopogon(Stapf)Koss。(4)星接藻科Zygnemataceae:水绵 Spirogyra communis(Hass.)Kutz。(5)泽泻科 Alismataceae:矮慈菇 Sagittaria pygmaea Miq.。(6)莎草科 Cyperaceae:碎米莎草Cyperus iria L.,咸草 Cyperus malaccensis Lam,异型莎草Cyperus difformis L.,扁秆藨草Scirpus planiculmis Fr.Schmidt,荸荠苗 Heleocharis dulcis Trin.ex Henschel。(7)浮萍科 Lemnaceae:紫萍 Spirodela polyrrhiza(L.)Schleid,青萍 Lemna minor L.。(8)灯心草科 Juncaceae:笄石菖 Juncus prismatocarpus Brown。(9)柳叶菜科Onagraceae:丁香蓼Ludwigia prostrata Roxb。(10)伞形科 Umbelliferae:水芹菜Oenanthe javanica(Blume)de Candolle。(11)玄参科Scrophulariaceae:陌上菜 Lindernia procumbens(Krock.)Borbas。(12)雨久花科 Pontederiaceae:鸭舌 草 Monochoria vaginalis(Burm.f.)Preslex Kunth,水葫芦 Eichhornia crassipes(Mart.)Solms。(13)蓼科 Polygonaceae:酸模 Rumex acetasa L.。(14)满江红科 Azollaceae:满江红 Azolla imbricata(Roxb.)Nakai。(15)苋科 Amaranthacae:空心莲子草 Alternanthera philoxeroides(Mart.)Griseb.。(16)菱科 Trapaceae:河菱 Trapa bicornis Osbeck。(17)菊科 Asteraceae:鳢肠 Eclipta prostrata L.。(18)天南星科 Araceae:菖蒲 Acorus calamus L.,芋头苗 Colocasia esculenta(L.)Schoot。(19)香蒲科Typhaceae:香蒲Typha orientalis Presl,小香蒲 Typha minima Funk。(20)睡莲科 Nymphaeaceae:藕 Nelumbo nucifera Gaertn。(21)旋花科Convolvulaceae:空心菜 Lpomoea aquatica Forsskal。(22)茄科 Solanaceae:辣椒 Capsicum frutescens L.。(23)葫芦科Cucurbitaceae:苦瓜 Momordica charantia L.。(24)十字花科Brassicaceae:白萝卜Raphanus sativus L.。
2.2 福寿螺的取食情况
因不同种样品植株在水中会有不同程度的生长及吸水增重,为尽量避免试验的系统误差,选用较为合理的受试植物(样品)相对减少率作为处理螺区的取食量衡量指标。福寿螺对植物(样品)啮食有趋细嫩的习性。在受试的寄主植物(样品)中,表皮角质柔软细嫩的满江红、紫萍、水葫芦、菹草、槐叶萍等5种植物在放入福寿螺1 d即被完全啮食;鸭舌草、碎米莎草、陌上菜、矮慈菇等4种植物在放入福寿螺2 d即被完全啮食;辣椒、青萍、水绵、稗草及苦瓜也在放入福寿螺3 d后被完全啮食。相对较难被啮食的如香蒲,放螺后3 d与放螺后5 d的相对取食率相差较少,而菖蒲、扁秆藨草、鳢肠在放螺后5 d相对取食率反而比放螺后3 d还低,推测其原因在于成螺不易取食较硬的样品,并在处理3 d后进入低食量期以适应环境,而处理的样品植株在经过前期的轻微危害后反而比对照生长得更快。个别植物如芋头苗、牛筋草等在放螺3 d后表现出较多的相对取食量,推测其原因在于这些植物浸泡3 d吸水软化后变得较易被福寿螺取食(表1)。
上述结果表明,福寿螺喜欢取食水生和湿生作物的绿色叶子或细嫩的芽,根、茎等老硬部分需在水中浸泡变软或腐烂后才可被取食;陆生植物(包括块根、块茎等)投入水中或经水浸泡后,也可被取食;刺激性较强的苦瓜、辣椒等植物的果实对其没有趋避性。
2.3 寄主植物对福寿螺体重的影响
福寿螺取食不同的受试植物(样品)后,体重增长率差异较大(表2)。满江红、紫萍、水葫芦、菹草、槐叶萍、陌上菜、矮慈菇等植物在放入福寿螺后因短期内就被大量取食,使其体重极显著增加;取食辣椒、苦瓜等食物的福寿螺体重增加也较显著,其中,辣椒、苦瓜、千金子、芋头苗、茭白苗等5种植物(样品)被取食5 d后,每天每只成螺的体重增长率在5%以上,有利于成螺的快速生长,从而提高了孕卵量和产卵量;取食满江红、水稻苗、水稗等19种植物5 d后,每天每只成螺的增长率为1%~5%;小香蒲、空心莲子草、荸荠苗、黑麦草、水芹菜、陌上菜、扁秆藨草、藕、稗草等9种植物被取食5 d后,每天每只成螺的增长率在1%以下;而菖蒲、水绵、鳢肠、咸草、酸模等5种植物尽管能被福寿螺取食,但取食5 d后,福寿螺体重几乎没有增加。

表1 福寿螺对受试植物(样品)啮食情况Table 1 Food consumption and relative feeding rate of GAS on different plants in the no-choice assays
2.4 取食率与生长率之间的关系
将相对取食率(x)与成螺增长率(y)做回归分析,得到:y=0.0241x+0.622,r=0.3538,df=32,P<0.05(图1)。由此可以初步得出结论:短期内的相对取食率对福寿螺的体重增长率有显著的影响,即取食越多生长越快。将相对取食率(x)和成螺增长率(y)均按最大和最小值对半划分,进一步将福寿螺的摄食植物分成4类,相当于二维平面坐标空间的4个象限:Ⅰ多食快长型,如千金子、异型莎草、满江红、辣椒及苦瓜;Ⅱ少食快长型,如茭白;Ⅲ少食慢长型,如水稻、菖蒲、水花生、黑麦草、香蒲、双穗雀稗等;Ⅳ多食慢长型,如稗草、陌上菜、水绵、河菱、藕、马唐、荸荠、青萍、水蓼(丁香蓼)、水芹菜、纤毛鹅观草、笄石菖、牛筋草、芦苇等(图1)。
2.5 植物总酚含量与福寿螺生长率之间的关系
将田迪英和杨荣华(2004)测定的41种果蔬中与本文重叠的6种植物的总酚含量数据,与本试验中福寿螺5 d的相对取食率、体重增长率做相关分析,发现5 d的相对取食率与总酚含量无显著相关性(r=0.2371,P >0.10);而5 d 的体重增长率与总酚含量之间呈显著的负相关:y=-1.0138x+6.3646,r=0.7465,P <0.10(图 2)。水芹菜和藕属于总酚含量高的植物(田迪英等,2004),虽取食率高但生长慢,即属于“Ⅳ型”;而苦瓜、茭白总酚含量较低,取食率高,生长也较快,即属于“Ⅰ型”。

表2 取食不同植物(样品)对福寿螺体重的影响Table 2 Body mass increase of GAS fed on different plants provided during the no-choice assays
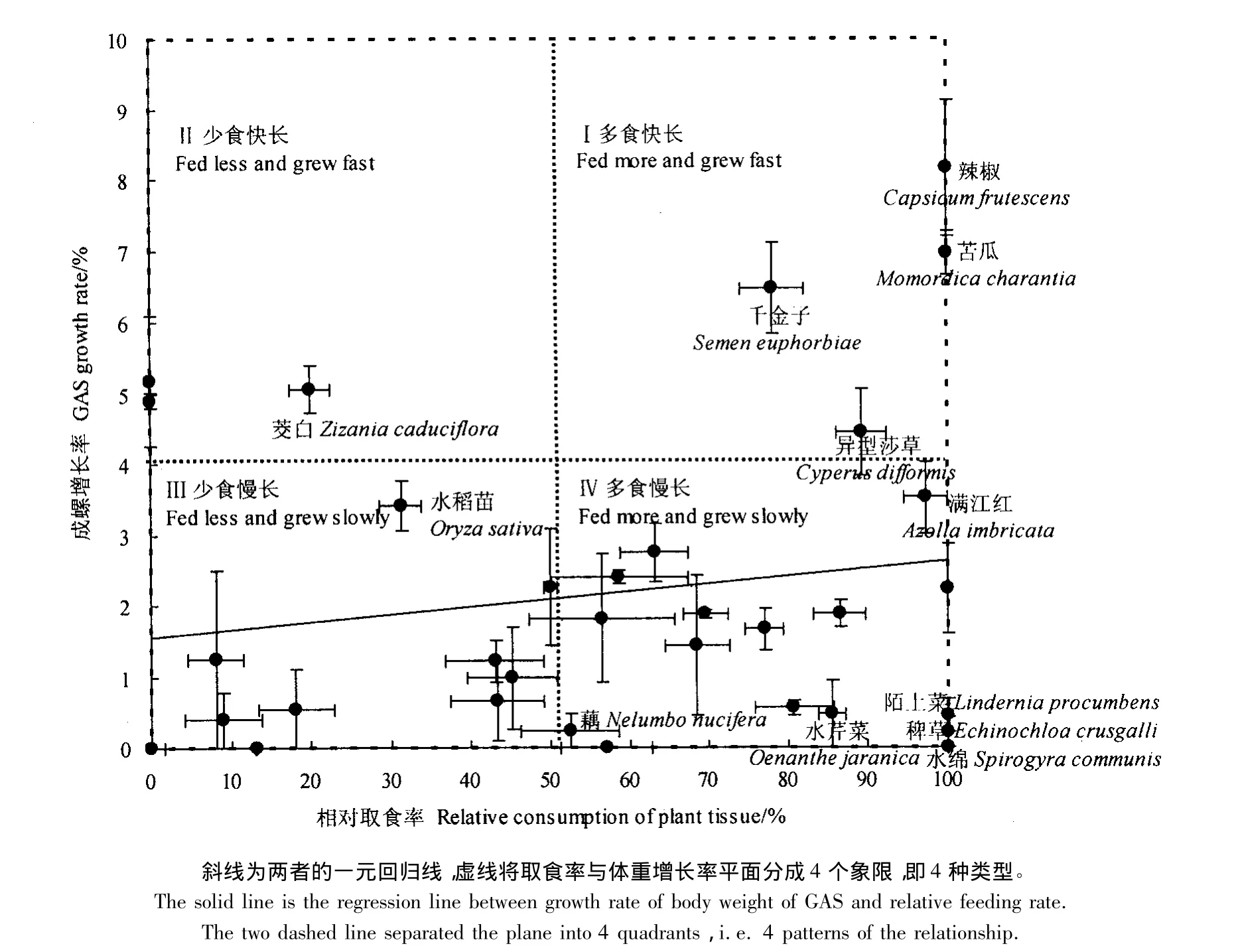
图1 福寿螺对植物的相对取食率与体重增长率之间的关系Fig.1 Relation of relative feeding rate of GAS on different plants to growth rate of body mass
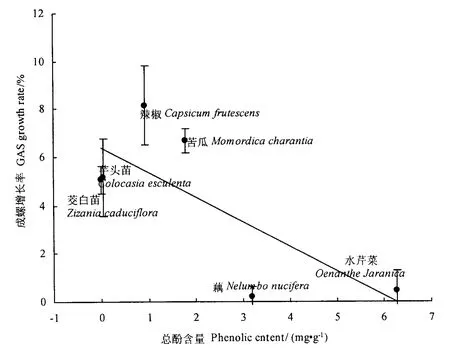
图2 6种植物的总酚含量与福寿螺生长率之间的关系Fig.2 Relation of growth rate of GAS to six host plants phenolic content
3 讨论
3.1 福寿螺的广食性是其快速扩散的重要原因
广食性动物(即多食性动物polyphagous animal)是指能以多科植物作为食物的动物。本试验结果表明,在32种常见的植物中,福寿螺对27种的相对取食量超过10%。可见,福寿螺是一种多食性的软体动物。同时,根据有关文献(Joshi&Sebastian,2006),福寿螺还可以攻击活的小动物,也能取食鱼类尸体,因此,它还是杂食性动物(omnivorous animal)。但一般仍以绿色植物为主,不仅取食水生或湿生植物的根、茎、叶以及块根、块茎等,而且取食掉落水中或受水浸泡的陆生植物的根、茎、叶以及块根、块茎等。食性范围广(韩雪梅等,2002),为其在新入侵区建立种群提供了必要条件,增加了福寿螺对环境的适应性。
本研究确立了水稻苗、菖蒲、异型莎草、菹草及苦瓜等稻田水域及周边广为分布的24科43种植物与福寿螺之间的营养关系。一些植物种(如鹅观草、菹草等)还属在国内的首次报道,且发现福寿螺在取食一些果蔬(如辣椒、苦瓜等)后体重较快增加,有利于其种群的建立和繁衍。而已知的几种有杀螺效果的植物(如夹竹桃、魔芋、乌药等)都不在水域生长(闫小红等,2011)。大部分水生植物对福寿螺的扩散有促进作用。
福寿螺不回避苦瓜、辣椒等陆生植物果实的苦辣味。我们认为,福寿螺的取食选择性不受其味觉影响,因此不能以苦、辣味植物作驱避植物来防治福寿螺的危害。这在一定程度上表明福寿螺的味觉与哺乳动物的味觉感受有很大的差异,有关福寿螺取食嗜好性的味觉、嗅觉机理有待进一步研究。
3.2 植物总酚含量是影响福寿螺生长的重要因子
酚类物质是植物防御植食者、病原菌取食危害的重要物质。含酚量低的鸭跖草科Commelinaceae裸花水竹叶Murdannia nudiflora的新鲜叶片含氮量高,被福寿螺取食的量大且有利其生长,而腐败的叶片含氮量低,不利其生长;小二仙草科 Haloragaceae粉绿狐尾藻Myriophyllum aquaticum的腐败叶片含氮量中等,含酚量低,氮和酚含量较高的新鲜叶片更易被福寿螺取食且利于其生长;蓼科Polygonaceae毛蓼Polygonum barbatum的新鲜和腐败叶片的含氮量均低,含酚量均中等,被福寿螺取食的量很小,且存活率低(Qiu et al.,2011)。总之,福寿螺取食含氮量适中且含酚量低的新鲜或腐败的植物更利于其生长。
本研究表明,植物的总酚含量不会影响福寿螺取食,但确实是影响其生长的重要因子。由于福寿螺为淡水螺类,主要入侵危害南方广阔的稻田生态系统,在该系统中像马齿苋、水芹菜、藕等含酚量高的植物丰盛度远不如水稻[三叶一心时叶片的总酚含量为 1.07 ~ 1.43 mg·g-1(鲜重)](周洁等,1997)、茭白[茎总酚含量为 0 mg·g-1(鲜重)](田迪英和杨荣华,2004)等含酚量低的作物类植物多,对福寿螺的总体生长抑制作用不明显,因此,福寿螺在南方水生生态系统中扩散十分迅速。但是,田迪英和杨荣华(2004)测定的这些植物主要为可食用部分的总酚含量,与本试验所用的植物叶片、茎秆不尽一致,但至少提供了一些进一步研究的线索,即测定本研究涉及的植物种类相应部分的总酚等次生化合物含量,深入了解福寿螺取食与其生长之间的关系(Li et al.,2009)。
3.3 借助福寿螺对植物的不同利用类型可防范其扩散和危害
本试验发现,福寿螺在取食率高的植物上相对生长率也高,不同的植物对福寿螺的生长有显著的影响。因此,将寄主植物分成4种类型:Ⅰ多食快长型,Ⅱ少食快长型,Ⅲ少食慢长型,Ⅳ多食慢长型;同时,福寿螺在较嫩的寄主植物上表现出“暴食”的现象,而在不宜取食的植物上则可以长期在低食量状态下正常生活,但长期低食量对成幼螺的生长存活有一定的影响(Qiu et al.,2011)。因此可以在福寿螺对植物的不同利用类型划分,以及比较精细的南方农田生态系统中不同类型的植物分布等景观成分组成的基础上,采用地理信息系统等时空分析技术,建立福寿螺的分布和扩散与这些景观成分之间的关系方程,预测福寿螺的分布和扩散,并通过生态设计提出景观层次的生态工程治理技术体系,以抑制福寿螺的扩张,有效防治其危害。对于少食快长型的茭白等作物,必须做好及时有效的防范;严禁将多食快长型的蔬果废弃物向水面倾倒;在福寿螺分布密集的河道附近种植多食慢长型的藕类作物,保留河道边的水芹菜等多食慢长型和一些少食慢长型杂草,作为生物屏障;福寿螺偏好取食植物的嫩芽新叶,如分蘖期以内的水稻嫩苗有利于福寿螺的取食和生长(Joshi&Sebastian,2006),当水稻成株时福寿螺对本季节水稻则不会造成直接危害(符明龙等,2008),因此,对于少食慢长型植物特别是水稻等重要作物,应在秧苗期加强对福寿螺的重点防治,在移栽时应尽量选择大苗壮苗。
尽管已报道的治理福寿螺的策略和措施很多,如使用杀螺剂等化学防治、养鸭和甲鱼等生物防治(陈建明等,2003)、使用防螺网等物理防治手段,但其仍日益扩散并有愈演愈烈的趋势。这不仅与本文指出的福寿螺食性广杂、取食率与生长呈正相关、水生生境中具备抑制作用的总酚含量高的植物丰盛度低等有关,而且与福寿螺的幼螺和成螺的主动和被动扩散能力密切相关(符明龙等,2008;叶建人和李云明,2009;叶建人等,2011),更重要的是人为携带(Joshi&Sebastian,2006)。因此,对福寿螺的已入侵区进行隔离并阻止其逃逸是重要的防扩散措施;同时应加强农民、消费者、宠物爱好者等对福寿螺危害性的认识。
陈建明,俞晓平,郑许松,徐红星,吕仲贤,张珏锋.2003.茭白田福寿螺的生物学特性和无害化治理技术.浙江农业学报,15(3):154 -160.
符明龙,谢淑玲,祝增荣,陈辉珍,王华弟.2008.入侵移栽稻田的福寿螺卵块空间分布型及其抽样技术.植物保护学报,35(4):332-338.
韩雪梅,徐岩,张清芬,宋红敏,徐汝梅.2002.134种植物害虫的寄主与扩散分析.植物检疫,16(6):331-335.
环保总局和中国科学院.2003.关于发布中国第一批外来入侵物种名单的通知(环发〔2003〕11 号).http://www.gov.cn/gongbao/content/2003/content_62285.htm.
唐启义.2010.DPS数据处理系统.北京:科学出版社.
田迪英,杨荣华.2004.果蔬抗氧化活性与总酚含量相关性研究.化学世界,(2):70-74.
万方浩,郭建英,张峰.2009.中国生物入侵研究.北京:科学出版社.
闫小红,周兵,邹有,蒋平,许冬梅.2011.16种植物提取物对福寿螺杀螺效果的研究.广东农业科学,(4):81-84.
杨叶欣,胡隐昌,李小慧,汪学杰,牟希东,宋红梅,王培欣,刘超,罗建仁.2010.福寿螺在中国的入侵历史、扩散规律和危害的调查分析.中国农学通报,26(5):245-250.
叶建人,李云明.2009.福寿螺在水稻田的消长规律及其田间分布动态.中国农学通报,25(3):185-188.
叶建人,张桂明,黄贤夫,蔡娟娟,陈海波,冯永斌.2011.稻鸭共育对水稻田福寿螺的控制作用.浙江农业科学,(1):114-116.
尹绍武,颜亨梅,王洪全.2000.福寿螺的生物学研究.湖南师范大学自然科学学报,23(2):76-82.
俞晓平,和田节,李中方,吕仲贤,孙乐平,朱亚红,陈建明,郑许松,徐红星.2001.稻田福寿螺的发生和治理.浙江农业学报,13(5):247-252.
章家恩,方丽.2008.关于我国农田福寿螺生物入侵需要加以研究的生态学问题.中国生态农业学报,16(6):1585-1589.
周洁,陈伟,叶明志,夏怡厚.1997.水稻感染细菌性条斑病后叶片中酚类物质的变化.福建农业大学学报,26(2):250-255.
Joshi R C and Sebastian L S.2006.Global Advances in Ecology and Management of Golden Apple Snails.Nueva Ecija:Phil-Rice.
Li K Y,Liu Z W,Hu Y H and Yang H W.2009.Snail herbivory on submerged macrophytes and nutrient release:implications for macrophyte management.Ecological Engineering,35:1664-1667.
Morrison W E and Hay M E.2011.Feeding and growth of native,invasive and non-invasive alien apple snails(Ampullariidae)in the United States:invasives eat more and grow more.Biological Invasions,13:945 -955.
Naylor R.1996.Invasions in agriculture:assessing the cost of golden apple snail in Asia.Ambio,25:443 -448.
Qiu J W,Chan M T,Kwong K L and Sun J.2011.Consumption,survival and growth in the invasive freshwater snail Pomacea canaliculata:does food freshness matter?.Journal of Molluscan Studies,77:189 -195.
