类芋螺毒素基因的克隆及其斜纹夜蛾重组杆状病毒转移载体的构建
2011-09-11孙兴鲁汤欣欣王文兵史全良浦冠勤
孙兴鲁 汤欣欣 朱 江 王文兵 史全良 浦冠勤
(1苏州大学金螳螂建筑与城市环境学院,江苏苏州 215123; 2苏州大学医学部基础医学与生物科学学院,江苏苏州 215123;3江苏大学生命科学学院,江苏镇江 212013)
芋螺毒素(Conotoxin,CTX)是由生活在热带海洋中的肉食性软体动物芋螺(Conus)分泌的,用于麻醉猎物的小肽类毒素。它们多数是由12~46个氨基酸残基组成,富含半胱氨酸(Cys)的动物神经肽毒素。芋螺毒素毒性极大,可引起动物出现惊厥、颤抖、甚至麻痹死亡[1]。
目前,国内外已明确功能的芋螺毒素只占了整个毒素库的很小一部分,但是对它们的生理功能已经有了较为清晰的认识。芋螺毒素二硫键骨架和结构的不同,决定了它们功能靶位的差异,已经清楚的靶位主要包括配体门控的离子通道、电压门控的离子通道及G蛋白偶联的受体[2]。芋螺毒素有如下特点:分子质量小,结构多样;前导肽高度保守而成熟肽具有超变异性;作用靶点广且具有高度组织选择性。因而,较之其他生物来源的活性多肽具有更多的优越性。
应用昆虫杆状病毒对害虫进行防治,是害虫综合防治的一项重要内容。1975年美国环保总局批准了第1个用于防治棉花害虫的美洲棉铃虫核型多角体病毒(Heliothis zeaNPV)杀虫剂产品登记[3]。但是,野生杆状病毒杀虫剂具有杀虫速度慢、寄主范围窄、对紫外线敏感等缺点,限制了其作为生物防治因子的广泛应用[4]。为了提高其杀虫效力,通常将野生的杆状病毒进行基因工程改造,包括删除、修饰昆虫病毒基因组内的某些非必需基因,如缺失蜕皮激素甾体-UDP 葡萄糖基转移酶基因(egt)[5-7]以及插入一些对昆虫特异性的毒素基因,调控昆虫代谢或发育的基因以及某些酶基因等外源基因,以构建重组病毒[8-12]。
斜纹夜蛾(Spodoptera litura)是一种多食性的重要农业和林业害虫,危害植物达99科290种以上[13],在中国、日本、朝鲜、印度以及东南亚地区经常造成严重危害和巨大经济损失[14-15]。斜纹夜蛾也是桑树的重要鳞翅目害虫之一。在我国,斜纹夜蛾核型多角体病毒(SpltMNPV)作为商品化杀虫剂已推广使用[16-17],但仍有野生杆状病毒杀虫剂的诸多不足。本研究以繁殖率和毒力极强且基因组中不含有类芋螺毒素成熟肽基因的病毒变异株斜纹夜蛾核型多角体病毒株SpltMNPVⅡ(GenBank:NC_011616)为基础,构建缺失egt基因,同时在缺失egt基因的部位插入多角体基因启动子控制的类芋螺毒素成熟肽基因和p10基因启动子启动的增强型绿色荧光蛋白基因(egfp)的重组载体,为构建和筛选高效重组SpltMNPV病毒,进一步研究与开发斜纹夜蛾生物杀虫剂奠定基础。
1 材料和方法
1.1 材料
SpltMNPVⅡ病毒株,由日本岐阜县生物产业技术研究所Katsumi Kamiya博士馈赠;斜纹夜蛾TUAT-Spli 221细胞,由日本名古屋大学资源昆虫研究室Michihiro Kobayashi教授馈赠。苜蓿银纹夜蛾核型多角体病毒(AcMNPV)病毒株和Sf9细胞,载体质粒pBluescript II SK(+)、转化用宿主菌E.coliTop10,由苏州大学基础医学与生命科学学院微生物学研究室保存。Spli细胞和Sf9细胞,用含10%胎牛血清(Invitrogen公司)的TC-100培养基(Invitrogen公司)28℃ 培养。pMD18-egfp,由广东海洋大学刘艳荷博士馈赠。Ex-Taq DNA聚合酶、限制性核酸内切酶、载体质粒pMD18-T载体、T4DNA连接酶等,均购自TaKaRa公司。
1.2 病毒感染及基因组DNA的提取
实验使用80 cm2(Corning公司)培养瓶培养的单层贴壁细胞(1×l06cells/mL),除去培养液后接种1 500μL AcMNPV或SpltMNPV病毒液,感染复数(MOI:multiplicity of infection)为1,室温下用震荡器慢摇1 h,使病毒吸附细胞,然后除去病毒接种液,加入13 mL TC-100培养基(含10%胎牛血清),28℃培养96 h后收取病毒感染的培养细胞。根据参考文献[18]的方法从感染的细胞中提取病毒DNA。
1.3 类芋螺毒素基因的分子克隆
参照GenBank登录的苜蓿银纹夜蛾核型多角体病毒(AcMNPV)基因组全序列(登录号:NC_001623)ORF3(Ac-ctx)读码框序列,设计一对引物(表1)。以AcMNPV基因组全序列为模板进行PCR扩增,扩增条件为94℃变性5 min,预变性后按94℃ 30 s,55℃ 30 s,72℃ 30 s,扩增30个循环,72℃延伸10 min后,4℃终止反应。DNA片段分离、连接转化、质粒制备与纯化、限制性核酸内切酶酶切,均按参考文献[19]的方法进行,构建的重组质粒命名为pMD18-ctx。

表1 用于PCR扩增的引物序列
1.4 重组转移载体的构建
根据GenBank SpltMNPVⅡ基因组全序列(NC_011616)分别设计egt上游非编码端(egt up),位于egt3'部分片段和egt下游非编码端(egtdown),多角体启动子(Pph),F蛋白(Fusion protein)的信号肽(Signal peptide)基因引物(表1)。以上引物由上海生工生物工程技术有限公司合成。DNA片段分离、连接转化、质粒制备与纯化、限制性酶切等分子克隆方法,均按参考文献[20]的方法进行。
重组转移载体的构建流程如图1。以SpltMN-PVⅡDNA为模板,分别扩增出同源重组臂egt上游非编码端(egtup)、egt3'端的部分片段和egt下游非编码端(egtdown)。PCR程序:94℃预变性3 min;94℃变性45 s;55℃退火45 s;72℃延伸90 s,30个循环;72℃再延伸10 min;4℃,10 min。扩增产物回收纯化后分别经KpnⅠ/XhoⅠ,NotⅠ/SacⅠ双酶切后,依次克隆到载体pBluescriptⅡSK(+)上。鉴定为阳性克隆且外源基因插入方向与预期一致后,命名为psk-egtup-egtdown。通过三引物PCR将多角体启动子基因序列与F蛋白信号肽序列连接,经T-A连接、转化获得pUC-Pph-signal P;经XhoⅠ/SmaⅠ双酶切,将其克隆至psk-egtup-egtdown上,获得psk-△egt-Pph-signal P;依次克隆Pp10与egfp到质粒pBacPAK8上,命名为pBacPAK8-Pp10-egfp;经BamHⅠ/NotⅠ双酶切,将其克隆到psk-△egt-Pph-signal P上,命名为psk-△egt-Pph-Pp10-egfp;最后经SmaⅠ/BamHⅠ双酶切pMD18-ctx,将其克隆到 psk-△egt-Pph-Pp10-egfp上,最终获得 psk-△egt-Pph-ctx-Pp10-egfp。委托上海生工生物工程技术有限公司进行序列测定。

图1 重组转移载体psk-△egt-P p h-ctx-P p10-egfp构建策略
2 结果与分析
2.1 AcMNPV类芋螺毒素基因(Ac-ctx-like)的克隆与序列分析
2.1.1 克隆基因Ac-ctx-like的鉴定 PCR扩增产物经1%琼脂糖凝胶电泳,在接近100 bp处可见特异性 DNA扩增带(图2)。回收 Ac-ctx-like的PCR产物,然后与pMD18-T载体连接、转化,提取质粒。经SmaⅠ和BamHⅠ双酶切鉴定,结果与预期大小一致,表明已成功克隆Ac-ctx-like基因(图3)。克隆基因委托上海生工生物工程技术有限公司进行序列测定。

图2 Ac-ctx-like基因的PCR产物琼脂糖凝胶电泳鉴定
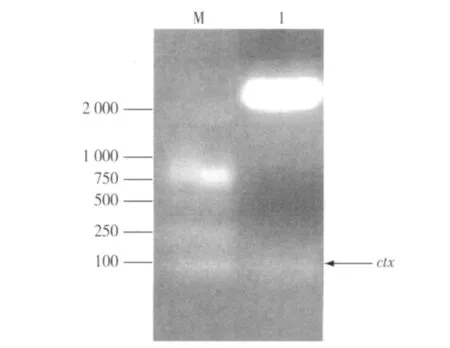
图3 pMD18-ctx双酶切的琼脂糖凝胶电泳鉴定
2.1.2 克隆基因Ac-ctx-like的序列分析 序列分析表明,Ac-ctx-like基因为162 bp,其中前72 bp为信号肽序列(图4)。为了更好地进行表达和分泌,本研究使用了斜纹夜蛾F蛋白的信号肽序列,并将该序列构建到转移载体上,所以克隆的Ac-ctx-like基因是从该基因的第25个氨基酸密码子开始的90 bp片段。

图4 Ac-ctx-like基因的DNA序和氨基酸序列
根据芋螺毒素成熟肽半胱氨酸残基间的二硫链键桥的特征性框架排列模式和芋螺毒素前体N端区域氨基酸的高度保守的信号肽序列,芋螺毒素可以分成A、M、O、P、S、T等多个超家族。又可按照其作用靶位的不同,分为 α、μ、ω、κ、δ、φ 、σ、ρ、γ 及加压素、惊厥剂和睡眠肽等。根据Ac-ctx-like基因推定的成熟肽半胱氨酸残基间的二硫链键桥的特征性框架排列模式(图5),推测该Ac-ctx-like基因可能为类ω-芋螺毒素基因。ω-芋螺毒素是电压敏感性钙通道(Voltage sensitive calcium channel,VSCC)阻滞剂,属于芋螺毒素O超家族[20]。

图5 ω-芋螺毒素成熟肽半胱氨酸残基间二硫链键桥的特征性框架排列模式
2.2 重组转移载体psk-△egt-P ph-ctx-P p10-egfp的构建与鉴定
2.2.1 Signal P+Pph基因片段的TP-PCR连接与鉴定 由于该重组转移载体的插入片段较多,为减少载体的限制性酶切位点,采用三引物PCR(TP-PCR)将Signal P基因与Pph基因启动子序列连接,琼脂糖凝胶电泳结果表明,连接成功(图6)。对目的基因片段回收,经连接、转化获得质粒pUC-ctx,委托上海生工生物工程技术有限公司进行序列测定,测序结果与预期一致。
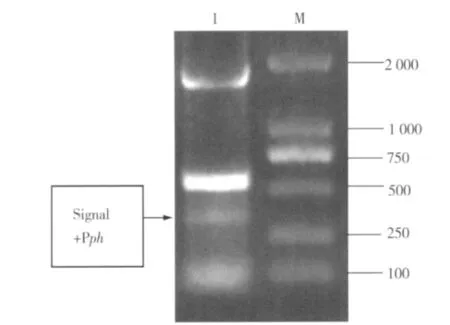
图6 Signal P+P ph基因片段琼脂糖凝胶电泳图
2.2.2 重组转移载体 psk-△egt-Pph-ctx-Pp10-egfp构建 重组转移载体构建各阶段均经限制性核酸内切酶酶切鉴定,结果正确(图7-8)。重组转移载体psk-△egt-Pph-ctx-Pp10-egfp经SmaⅠ /BamHⅠ双酶切鉴定,ctx片段大小与预期一致(图9)。经测序证明,重组转移载体包含了预期的各DNA片段,且克隆方向正确,无突变,上述结果证明该重组转移载体构建成功。
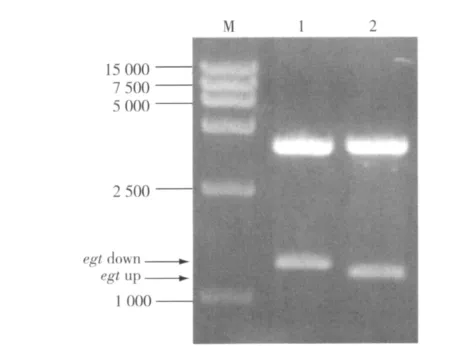
图7 重组转移载体egt up和egt down片段的双酶切琼脂糖凝胶电泳鉴定

图8 重组转移载体P p10+egfp片段的双酶切琼脂糖凝胶电泳鉴定
图9 重组转移载体psk-△egt-P ph-ctx-P p10-egfp的双酶切鉴定
3 讨论
SpltMNPVⅡ(SpltMNPV G10-3)是 Kamiya等从日本全国各地野外感病幼虫收集、分离到189个SpltMNPV克隆中,筛选出的繁殖率和毒力极强的一种新型病毒株[21-22]。本研究是在 SpltMNPV Ⅱ基础上,利用egt基因同源重组臂DNA序列,成功构建了含有多角体基因启动子启动的类芋螺毒素基因和p10启动子启动的增强型绿色荧光蛋白(egfp)基因的重组移载体psk-△egt-Pph-ctx-Pp10-egfp。为了使外源基因获得高效表达,在构建重组转移载体时,本研究使用了杆状病毒的强启动子——SpltMNPV的多角体基因启动子(Pph),并通过另一个杆状病毒的强启动子——p10启动子,启动增强型绿色荧光蛋白作为标记,这样可大大减少重组病毒的筛选与纯化的工作量和时间。本研究在ctx基因前引入了AcMNPV BV膜融合蛋白GP64的同系物,SpltMNPVⅡF(Fusion)蛋白的信号肽序列,可能有利于类芋螺毒素更好地分泌表达。用构建的重组转移载体与SpltMNPVⅡ在斜纹夜蛾细胞中共转染,筛选重组杆状病毒的工作正在进行中。
现已发现的芋螺毒素7大超家族,能特异性地作用于体内钾、钠、钙等多种离子通道、细胞膜上的各种神经递质和激肽的受体,从而干扰细胞或神经中的信号传递,有很强的生物学活性[23]。芋螺毒素目前尚无应用于重组杆状病毒研究的报道,本研究首次从AcMNPV中克隆了昆虫杆状病毒基因组中的类芋螺毒素基因,根据克隆基因的DNA序列及其推定的氨基酸序列的结构分析,推测该基因的表达产物可能类似芋螺毒素O超家族的类ω-芋螺毒素。
昆虫杆状病毒作为无公害生物杀虫剂近年来备受关注,并得到广泛应用。重组杆状病毒的研究已成为生物杀虫剂研究的主要方向。本研究结果将为进一步研制重组斜纹夜蛾核型多角体病毒生物杀虫剂打下基础。
[1]罗素兰,张本,长孙东亭.芋螺毒素[J].生物学通报,2003,38(4):7-8.
[2]吴雪晨,彭灿.芋螺毒素国内外研究现状[J].科技资讯,2007,(10):209-210.
[3]Inceoglu A B,Kamita SG,Hinton A C,et al.Recombinant baculoviruses for insect control[J].Pest Management Science,2001,57:981-987.
[4]Hu Z,Vlak J M.Engineering of biosafe baculoviruses with improved insecticidal properties:development and prospects[J].Virologica Sinica,1997,12:14 -25.
[5]Pinedo F JR,Moscardi F,Luque T,et al.Inactivation of the ecdysteroid UDP-glucosyltransferase(egt)gene ofAnticarsia gemmatalisnucleopolyhedrovirus(AgMNPV)improves its virulence towards its insect host[J].Biological Control,2003,27:336 - 344.
[6]O'Reilly D R,Miller L K,Luckow V A.A baculovirus blocks insect molting by producing ecdysteroid UDP-glucosyl transferase[J].Science,1989,245:1 110 -1 112.
[7]Georgievska L,Joosten N,Hoover K,et al.Effects of single and mixed infections with wild type and genetically modified Helicoverpa armigera nucleopolyhedrovirus on movement behaviour of cotton bollworm larvae[J].Entomologia Experimentalis Et Applicata,2010,135:56 -67.
[8]Maeda S.Further development of recombinant baculovirus insecticides[J].Current Opinion in Biotechnology,1995,6(3):313 -319.
[9]Miller L K.Genetically engineered insect virus pesticides:Present and future[J].Invertebr Pathol,1995,65(3):211 - 216.
[10]Jinn T R,Tu W C,Lu C I,et al.Enhancing insecticidal efficacy of baculovirus by early expressing an insect neurotoxin,LqhIT2,in infected Trichoplusia ni larvae[J].Applied microbiology and biotechnology,2006,72(6):1 247 -1 253.
[11]曹建斌,范晓军,付月君,等.重组杆状病毒AcMNPV-BmK IT-vcath的构建及毒力分析[J].山西大学学报:自然科学版,2009,32(1):114 -118.
[12]Shao H,Dong D,Hu J,et al.Construction of the recombinant baculovirus AcMNPV with cathepsin B-like proteinase and its insecticidal activity against Helicoverpa armigera[J].Pesticide Biochemistry and Physiology,2008,91(3):141 -146.
[13]梁东瑞,胡远扬,蔡毓能,等.中国昆虫病毒图谱[M].长沙:湖南科学技术出版社,1986.
[14]Murata M,Etoh T,Hoyama K,et al.Sudden occurrence of the common cutworm,Spodoptera litura(Lepidoptera:Noctuidae)in southern Japan during the typhoon season[J].Applied entomology and zoology,1998,33:419 -427.
[15]Gabriel B P.Insect and mites injurious to Philippine Crop Plants[R].UPLB:National Crop Protection Center,1997.
[16]苏志坚,陈其津,李广宏,等.斜纹夜蛾核多角体病毒与增效剂混配对斜纹夜蛾幼虫的防治效果[J].中国生物防治,2001,17(1):23-25.
[17]黄冠辉,丁翠.斜纹夜蛾核型多角体病毒的研究[J].昆虫学报,1975,18(1):17-23.
[18]Wu F,Lavina B,Iked A M,et a1.Cloning and biological characterization of Spod optera litura(Lepidoptera:Noctuidae)in nucleopolyhedrovirus isolated from China[J].Journal of Sericultural Science of Japan,2000,69(3):177 -189.
[19]Sambrook J,Fritsch E,Manlatis T.Molecular Cloning:A Laboratory Manual[M].New York:Cold Spring Harbor Laboratory Press,1989.
[20]应士波,严小军.芋螺毒素的药用价值研发进展[J].天然产物研究与开发,2006,18(1):138 -143.
[21]Kamiya K,Zhu J,Murata M,et al.Cloning and comparative characterization of three distinct nucleopolyhedrovirnses isolated from the common cutworm,spodoptera litura(Lepidoptera:Noctuidae)in Japan[J].Biological Control,2004,31(1):38 -48.
[22]朱江,沈颂东,王文兵,等.日本三株斜纹夜蛾核型多角体病毒的增殖特性及其多角体蛋白基因的序列分析[J].昆虫学报,2004,47(5):543 -550.
[23]王承忠,蒋辉,戚正武.芋螺毒素研究进展[J].生物化学与生物物理进展,2003,30(4):537 -543.
