流域植物生长与营养物质迁移转化模式研究
2011-08-29张万顺杨寅群仲兆林
张万顺,杨寅群,吴 静,仲兆林
(1.武汉大学 资源与环境科学学院,湖北 武汉 430079;2.江苏省水文水资源勘测局常州分局,江苏 常州 213001)
1 研究背景
流域内植物生长与土壤养分相互影响,相互作用。植物对土壤养分的吸收是流域营养物质循环的重要环节,土壤养分又是影响植物生长最主要的环境因素之一。植物在营养物循环中起到转化器的作用:植物从土壤中吸收的营养物质一部分经凋落物分解和秸秆还田等回归土壤中,一部分由于收割流出土壤-植被-水体组成的流域系统。因此流域营养物质迁移转化过程与植物生长有密切关系,而氮、磷等营养物质又是造成水体富营养化的主要污染物质。
植物-土壤系统中营养物质迁移转化的研究已有较长历史。20世纪70年代建立了两类系统模拟模型:一类是以土壤水分/营养物运移及转化利用为中心的模型,以美国的HYDRUSID[1]模型为代表,但它没有将植物生长过程考虑在内;另一类是以作物生长发育为中心的模型,如PS123模型[2],研究作物生长发育和耕作栽培措施,主要是土壤水分和养分条件对作物生长的影响,但对水分及营养物与作物生长关系的主要环节—根系生长和根系对水分和营养物吸收机理的处理尚需进一步完善。黄元仿等[3]建立了小麦、玉米地水氮运移的模型。张瑜芳等[4]建立了稻田土壤氮素的转化迁移模型。胡克林[5]等在前人研究成果的基础上,构建了土壤-作物系统农田水氮运移及作物生长联合模拟模型SPWS实现土壤氮素迁移转化的模拟。彭虹等[6]对汉江中下游水华问题以及张万顺[7]对汉江中下游水量水质耦合配置等研究中都考虑了植物生长对营养元素迁移转化的影响。
为反映流域内植物从生长直至死亡的过程对物质循环的影响,采用植物生长动力学模型,模拟生长过程不同阶段对水分及营养物质的吸收。模型将植物生长模型与土壤营养物质迁移转化模型耦合,详细模拟流域内生物量随时间变化和营养物物质迁移转化的动态变化过程,为土壤水水质和地表水面源污染物负荷的预测提供准确依据。
2 植物生长模型
为揭示流域水质变化机理,反映植物对营养物质转化的富集和吸收作用,确定流域内营养物质输入、输出和植物生长之间的耦合关系,开展植物生长模型研究。
从生物学角度研究植物生长模型较多,包括简单的单变量方程到植物生物量形成过程的复杂动力学模型[8-10]。但生物学模型多数只重视个别环境因子的作用和植物对资源的竞争和植物自身的变化,较少考虑到环境因子的多维性,无法全面反映流域系统中复杂外环境因素对植物生长的影响,特别是土壤水分与养分等外环境因素与植物生长过程的耦合关系。研究通过统计分析,从宏观上剥离影响植物生长的因素,得出植物理想状态下的最大生长速率,分别考虑温度、水分和养分3个主要外环境因素对植物生长的影响,求得多种外环境因素作用下的综合限制因子,模拟植物实际生长与外环境相互作用的动态过程。
2.1 理想生长温度、水分和养分等植物生长影响因子均处于最适宜状态时植物的生长速率为其最大生长速率。植物的最大生长速率在不同阶段是变化的,其值可由理想生长曲线得到。参考植物生长的相关研究文献[11-15],点绘出不同生长期对应的生物量。植物理想生长曲线为不同外部条件下不同时期累积生物量的外包线,如图1所示。对理想生长曲线求导得到最大生长速率变化过程线。研究划分的计算单元内植物类型通常有多种,单元内生物量的变化过程需依据耕作方式及单元内不同植物的种植面积占总面积的权重确定。
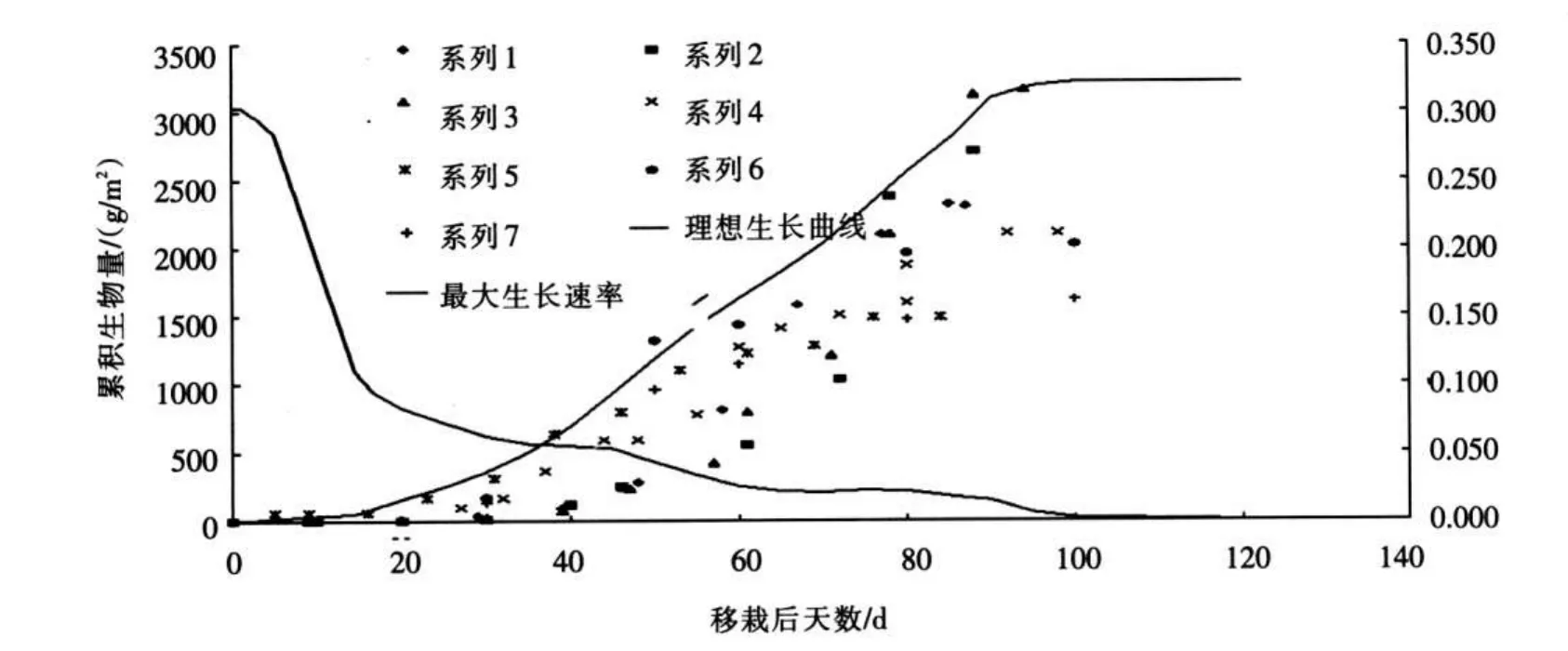
图1 典型植物理想生长曲线及最大生长速率
2.2 限制因子植物的实际生长会受到众多环境因素的影响,使得其生长速率小于理想状态下的最大生长速率。模型综合考虑温度、养分和水分的影响,计算植物不同阶段的实际生物量和营养物质吸收量。
2.2.1 温度因子 温度是控制植物生长的最重要的因素之一,每种植物的生长都需要一定的温度范围:生长的最低温度、适宜温度和最高温度。低于最低温度,植物停止生长;最低温度与理想温度之间,植物的生长速度随温度的升高而加快;高于适宜温度,植物的生长速度随温度的升高而降低,直至到最高温度生长速度降为0。温度限制因子为日平均气温与理想气温、最低气温及最高气温的函数,由下式计算:

式中:G(T )为植物生长的气温约束;T为日平均气温;Tbase为植物生长需要的基本或最低温度;Topt为植物生长的适宜温度;Thigh为植物生长所能耐受的最高温度。
通过对1961—2009年共39年典型流域逐日气温进行统计分析,以多年逐日平均气温作为适宜温度。调查不同植物生长期内各阶段所需基本温度和所能耐受的最高温度,并对生长期有重叠的各类主要植物的温度上限和下限进行平均化处理,分别得出温度上限和下限过程线。不同种类植物控制温度过程线以播种期为起点,当累积积温达到植物成熟所需值时为终点。太湖典型流域主要作物生长所需温度过程如图2所示。

图2 太湖流域植物生长温度过程线
2.2.2 养分因子 养分限值因子由植物在单位时间内营养物质的实际吸收量与理想吸收量的关系得到:
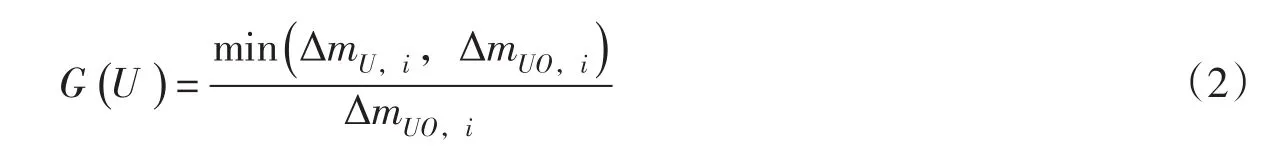
式中:ΔmU,i为第i天里植物的实际吸收营养物量(g);ΔmUO,i为第i天里植物的理想吸收营养物量(g)。
ΔmU,i由植物实际蒸腾量和土壤中溶解态(水相)营养物浓度求得:

式中:A为计算区域面积(m2);EC(i)为第i天的植物实际蒸腾量(mm);CU(i)为第i天时土壤中溶解态营养物的浓度(mg/l);ΔmUO,i由植物理想生长曲线和植物不同生长阶段植株相应营养物质含量占总生物量的比例求得。
植物生长需要多种养分,任何一种营养物质的缺乏都将影响植物的正常生长。模型考虑TN、NH3-N、TP及有机碳等4种对植物生长起主要作用的营养物质,以综合养分限制因子表征这4种营养物对植物生长的影响:

2.2.3 水分因子 水分限制因子为植物实际蒸腾量和最大蒸腾量的比值:

式中:Ec为植物实际蒸腾量(mm);Ecm为植物最大蒸腾量(mm),由Penman-Monteith公式算得。
Ec由下式计算:

式中:Droot是根系深度(cm);lroot(z )为根系在深度为z时的根系密度(cm·cm-3);αrw(h)是关于土壤水压力水头的一个无量纲函数,见图3。
αrw(h )值的变化范围为0~1。当αrw(h)等于1时,土壤根系吸水量等于潜在根系吸水量。当在土壤水压力水头大于厌氧点hl时,由于土壤中过多的水分导致根区缺氧,植物根系不能吸收水分;而当土壤压力水头低于凋萎点h4时,由于土壤太干,植物根系亦无法吸收水分;当压力水头在h2-h3之间时,植物根系可充分吸收水分,不会受到水分胁迫。

图3 根系吸水折减吸水与土壤水压力水头的关系
2.3 实际生长植物实际生长速率等于理想状态下的最大生长速率乘以综合限制因子,综合限制因子为温度、养分、水分限制因子的乘积。植物生长动力学模型需满足如下方程[8]:

式中:B为干重(g/m2);Gbmax(t)为植物最大生长率;G(T)为温度限制因子;G(NU)为营养限制因子;G(W)为水分限制因子。
2.4 叶面积指数与根系生长模拟由于植物的叶片和根系等器官对营养物质迁移转化有直接影响,在进行总生物量的模拟后,分别建立叶面积指数和根系生长模型。
2.4.1 叶面积指数模拟 叶面积指数(LAI)直接影响冠层截留和蒸散发等水循环过程主要环节,进而影响到与水循环密切相关的营养物迁移转化过程。在植物生长的初始阶段,叶面积由理想叶面积曲线确定。

式中: frLmx为给定积温条件下的最大叶面积指数的比例;ℓ1、ℓ2为形状系数; frp为计算时段内的积温占整个生长季节积温的比例。
生长期中植物叶面积指数随季节的变化情况如图4所示。
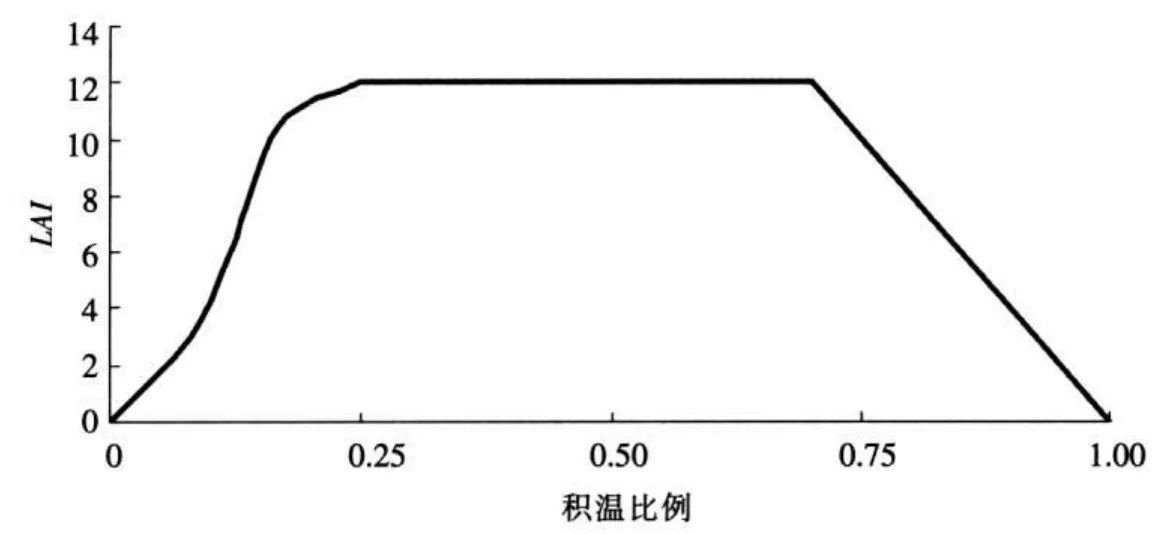
图4 生长期中植物叶面积指数随季节的变化情况
2.4.2 根系生长模拟 根系的深度和根系密度直接影响到植物对土壤水分的吸收,因此需对根系的生长情况进行模拟。根系生物量在播种阶段占总生物量的30%~50%,到成熟季节则下降到5%~20%。模型模拟时按照根系生物量在初始时占总生物量的40%,成熟后占总生物量的20%计算,且根系生物量与累积积温有关。根系生物量占总生物量的比例表示为:

则根系生物量为:

3 营养物迁移转化模型
3.1 流域营养物质循环机理模型模拟不同形态氮的迁移转化过程,包括地表径流流失、入渗淋失和化学输入等物理过程;有机氮矿化、反硝化等化学过程以及植物吸收等生物过程,如图5。
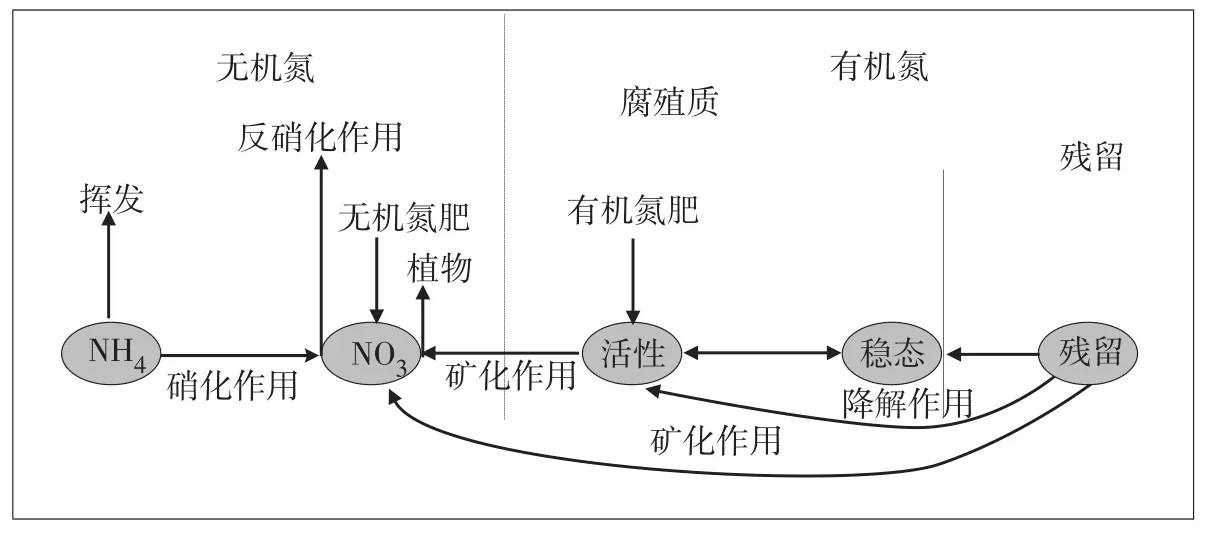
图5 模型模拟氮循环
磷可以分为腐殖质中的有机磷、不可溶解的无机磷和植物可利用的土壤溶液中的磷三种化学状态。磷可以通过施肥和残余物施用等方式添加到土壤中,通过植物吸收和径流携带从土壤中移除。磷可以与其他离子结合形成一些不可溶化合物,并从溶液中沉淀,使得磷在土壤表面累积,从而易于随地表径流运移,如图6。

图6 模型模拟磷循环
3.2 土壤溶质运移模型土壤中溶质处在物理、化学和生物的相互联系和连续变化的复杂运动中,不仅随着土壤水分的运动而迁移,还会在自身浓度梯度作用下运动,部分溶质可以被土壤吸附、为植物吸收或者当浓度超过水的溶解度后会离析沉淀,还有化合分解、离子交换等化学变化。土壤表面的不同的点位或位置,溶质的吸附、交换等物理化学特征不尽相同。充分考虑扩散、吸附解吸和微生物降解等情况,采用双点平衡/动力学吸附溶质运移模型,其控制方程为:
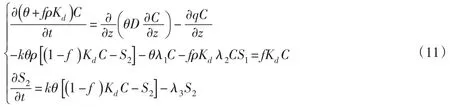
式中:C为污染物在水相中的浓度,mg/l;S1、S2分别为污染物在平衡和非平衡吸附相上的吸附浓度,mg/g;θ为体积含水率,%;f为平衡吸附交换点位占总吸附点位的比例,%; ρ为土壤体积密度,g/cm3;Kd为土壤-水分配系数,cm3/g;k为一阶吸附解吸速率常数;D为弥散系数,m2/d;λ1、λ2和λ3分别为水相、平衡吸附相和非平衡吸附相的一阶微生物降解速率系数,d-1。
4 模型在太湖流域典型区域的应用
4.1 研究区域概况太湖流域典型区位于江苏省南部,长江三角洲太湖平原西北部滆湖小流域。该区域为平原水网地区,区内大量圩区地势低洼,境内河流纵横密布,土壤水与地表水交换频繁;种植的农作物种类繁多,化肥农药使用量超过全国平均水平,居于全国前列,农业面源污染严重。
按照行政区域和水系特征的完整性、地表覆盖和土地利用、水功能区划等,研究区域划分为127个单元,选取75#单元体作为特征单元体,模拟典型作物种植情况下N、P的输出量。特征单元体位于常州境内,西北邻滆湖(入滆湖有一水质监测点),北傍太滆运河,南于宜兴相接,东与丹阳相连。单元内水稻种植占70%,棉花种植占20%,其他经济作物、小麦占10%。如图7所示。
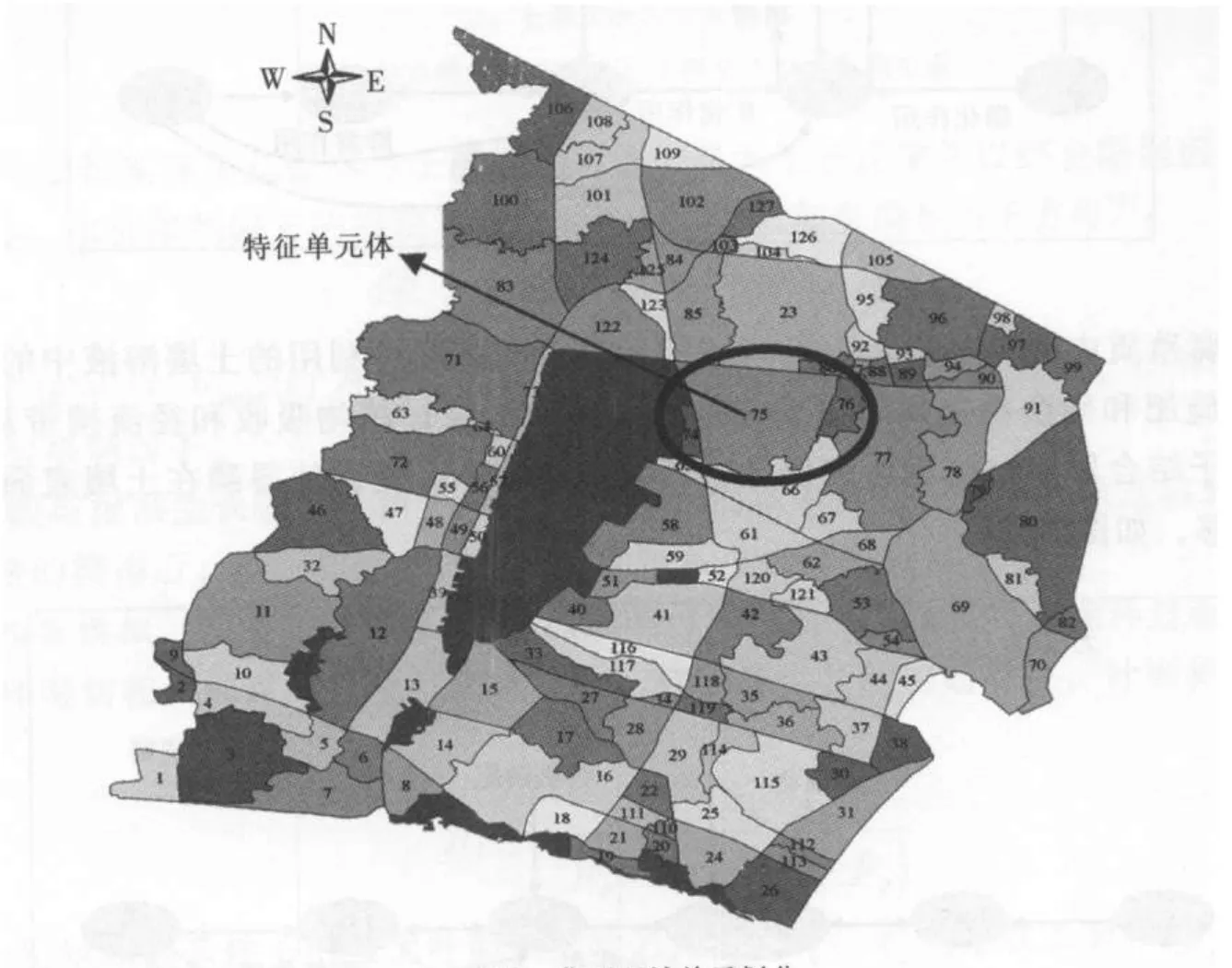
图7 典型区域单元划分
4.2 研究区域施肥特点太湖流域近年来施肥过程特点见表1[16]。

表1 太湖流域施肥过程特点 (单位:kg/hm2)
4.3 研究区域水文特征研究区降水量的年际变化幅度较大,多年平均降水量1 124mm,区域分布由南向北递减,梅雨期为6—7月,多年平均水面蒸发量为1 573.7mm,呈自南向北递增。采用泰森多边形法将流域内2010年点雨量转成面雨量,其年降雨量见图8。
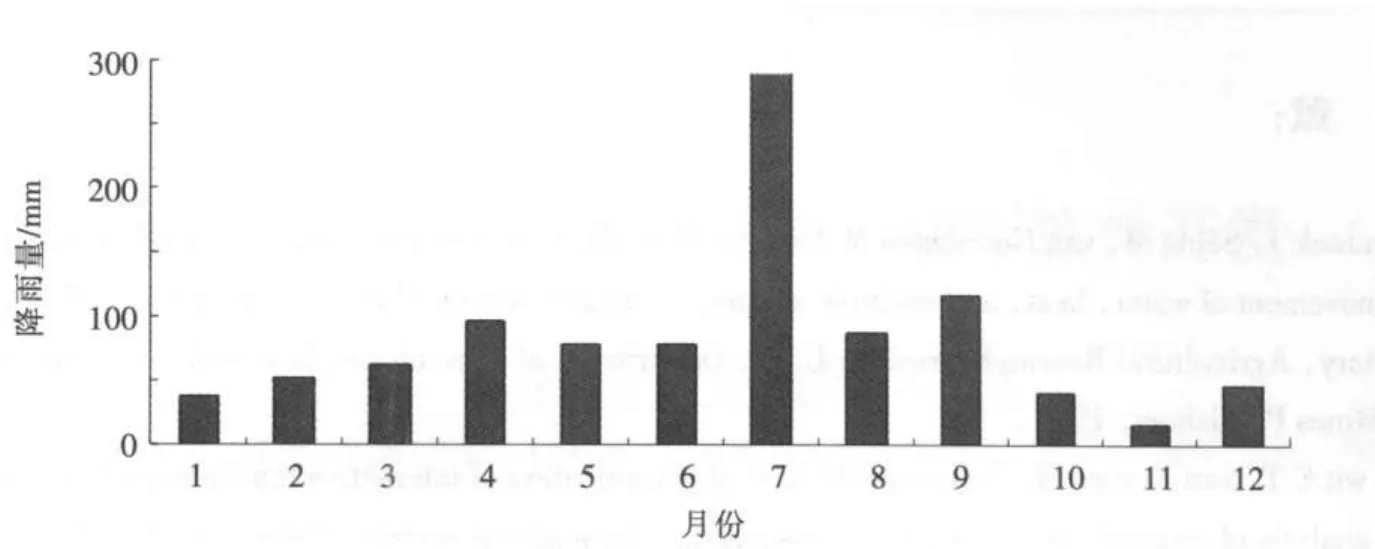
图8 特征单元体年降雨量
4.4 特征单元不同作物的污染负荷输出
(1)取特征单元出口入滆湖水样进行分析,将监测值与模拟值比较,见表2。

表2 特征单元营养物质N、P监测值与模拟值比较 (单位:mg/l)
由表2可以看出,监测值与模拟值基本相符。其中总氮的监测值在4月和8月与模拟值相差较大(由施肥期无准确时间造成),其他时期和总磷的值吻合较好。因此,采用本文提出的考虑农业施肥、植物生长对面源污染影响的方法是具有一定的精度的。
(2)不同作物对特征单元污染负荷输出的影响。净种水稻、小麦、棉花时,在作物生长过程中年输出总氮、总磷输出的模拟结果如表3。
由表3可以看出,根据本区域降雨特点,结合作物生长过程和施肥规律,特征单元体种植小麦在本单元体内输出的污染物最小,其中总氮为138kg/hm2,总磷为70kg/hm2。种植棉花则营养物质输出最大,总氮为287kg/hm2,总磷为101kg/hm2。

表3 植物生长对污染物输出的影响(单位:kg/hm2)
5 结论
(1)对特征单元,利用2010年的年降雨资料分析计算了种植棉花、小麦和水稻年总氮总磷的输出量,本单元在2010年降雨条件下种植小麦营养物质输出最小,总氮为138kg/hm2,总磷为70kg/hm2。而种植棉花营养物质输出最大,总氮为287kg/hm2,总磷为101kg/hm2。
(2)太湖地区氮污染较磷污染严重,约为磷污染的3倍。这与太湖地区氮肥施用过多有关,太湖地区氮肥施用量大约为磷肥施用量的3倍。在水稻生长季节,施氮肥总量为200kg/hm2,施磷肥总量为80kg/hm2;在小麦生长季节,施氮肥总量为225kg/hm2,施磷肥总量为90kg/hm2;在棉花生长季节,施氮肥总量为258.9kg/hm2,施磷肥总量为52.5kg/hm2。
(3)在三种作物生长过程中,小麦对环境污染最小,输出总氮为138kg/hm2,总磷为70kg/hm2;水稻次之,输出总氮为228kg/hm2,总磷为89kg/hm2;棉花对环境污染最大,输出总氮为287kg/hm2,总磷为101kg/hm2。作物生长对环境污染影响:棉花>水稻>小麦。
(4)在农业生产活动较多的月份,监测到的氮磷负荷明显增大。说明农业活动是环境氮磷污染的主要源头,是太湖流域面源污染物的主要来源。
[1]Simunek J,Sejna M,van Genuchten M Th.The HYDRUS-1D software package for simulating the one dimensional movement of water,heat,and multiple solutes in variably saturated media.Version 2.0[M]//U.S.Salinity Laboratory,Agricultural Research Service,U.S.Department of Agriculture,Riverside,California:Colorado School of Mines Publishers,1998.
[2]De wit C T,van Keulen H,Seligman H G,et al.Application of interactive multiple goal programming techniques for analysis of regional agricultural development[J].Agricultural system,1988,26:211-230.
[3]黄元仿,李韵珠,陆锦文.田间条件下土壤运移的模拟1-模型[J].水利学报,1996(6):9-14.
[4]张瑜芳,张蔚榛,沈荣开,等.排水农田中氮素转化运移和流失[M].武汉:中国地质大学出版社,1997.
[5]胡克林,李保国,陈研,等.作物生长与土壤水氮运移联合模拟的研究1-模型[J].水利学报,2007,38(7):779-785.
[6]彭虹,郭生练.汉江下游河段水质生态模型及数值模拟[J].长江流域资源与环境,2002,11(4):363-369.
[7]张万顺,方攀,鞠美琴,等.流域水量水质耦合水资源配置[J].武汉大学学报(工学版),2009,42(5):577-581.
[8]Gates D J.Competition between two types of plants located at random on a lattice[J].Math Biosci,1980,48(3):157-194.
[9]McMartrie R,Wolf L.A model of competition between trees and grass for radiation,water and nutrients[J].Ann Bot,1983,52(4):449-458.
[10]Walker B H,Ludwig D,Holling C S,et al.Stability of semi arid savanna grazing systems[J].J.Ecol,1981,69:473-498.
[11]彭世彰,李荣超,朱成立.节水灌溉的水稻干物质增长模型研究[J].水利学报,2002(11):99-102.
[12]田杰.稻草还田对土壤肥力及水稻生长的影响[J].安徽农学通报,2008,14(3):46-47.
[13]许显滨,横沢正幸,矫江,等.气温对水稻生育模型的影响研究[J].黑龙江农业科学,2005(2):5-7.
[14]孟亚丽,曹卫星,刘新伟,等.水稻光合生产与干物质累积的动态模拟[J].生物数学学报,2004,19(2):205-212.
[15]张俊平,陈常铭.水稻群体生长过程和产量的动态模拟[J].生态学报,1990,10(4):311-316.
[16]王鹏.基于数字流域系统的平原河网区非点源污染模型研究与应用[D].南京:河海大学,2006.
