蝴蝶多样性与地形相关性研究——以大峪流域为例*
2011-07-07张宇军房丽君
张宇军,房丽君
(陕西省植物研究所,陕西西安 710061)
蝴蝶物种多样性及其影响因素对蝴蝶资源调查与保护研究具有重要意义,气候等环境因子的变化对生物在海拔梯度上的影响越来越受到重视[1],海拔和物种丰富度呈显著的负相关关系[2]。一般认为,随着海拔的上升,热量的递减,物种多样性降低[3-4],在海拔梯度上,气候、环境变量在较小的空间范围内急剧变化,因此,物种丰富度沿海拔梯度的分布格局得到很高的关注[5],并且物种丰富度的海拔分布格局更加有利于探索影响物种丰富度的机制[6]。
地形在生态学中被看作间接的环境因子[7-8],但间接生态因子并不意味着是次要因子,相反,如果没有地形因子的差异,也就不会导致直接生态因子的重新分配,从而产生物种分布的多样性,因此,地形信息是生物多样性格局研究中不可缺少的变量,其中海拔被认为是物种多样性分布格局的决定性因素之一[9-11]。
数字高程模型简称DEM(Digital elevation model),它是用一组有序数值阵列形式表示地面高程的一种实体地面模型,具有形象、直观、精确等特点。GIS的空间分析功能是在DEM的基础上进行的,它能派生出各种地形因子,如坡度、坡向、平面凸率、剖面曲率、汇水面积等。它是进行地形分析的基础,广泛应用于测绘、遥感、地质、制图综合、地理形态、林业、资源、环境等领域[12]。
1 研究区域概况
研究区域为大峪流域,位于陕西长安县,属秦岭北坡,位于东经 109°06′55″~ 109°08′09″,北纬33°52′22″~ 34°01′03″之间,海拔 700 ~2 400 m,南北走向,中间有大峪河蜿蜒流出(图1)。植被以暖温带阔叶林为主,研究区内年均温为14~16℃,最冷月均温在0℃左右,最热月均温在20~26℃,无霜期200 d左右,年降水量800~900 mm,夏秋雨水丰沛,约占全年降水70%~80%。

图1 大峪流域地形模拟图
2 资料来源与数据分析方法
2.1 蝴蝶标本的采集
2009~2010年的5~8月期间在大峪流域进行实地采集调查,分类鉴定。建立包括科、属、种、沿海拔分布的蝴蝶物种分布数据库。
2.2 研究区域地形数据分析
本文采用的地形数据是ASTER GDEM(先进星载热发射和反发射仪全球数字高程模型)的30m分辨率的数字高程模型(DEM)及由此产生数字地面模型(DTM),用于提取大峪流域各种地形参数,利用地理信息系统软件A rcGIS提取海拔及面积等数据。
根据采集海拔高度将大峪流域从低海拔到高海拔按100 m为间隔划分为12个段,利用DEM数据,应用GIS软件计算每一海拔段的面积,平均海拔和海拔方差。为了表征不同海拔段的地形复杂程度,利用各海拔段的变异系数(cv)作为地形复杂度指数[13],用下式计算。地形复杂度(cv)=海拔方差/该海拔段平均海拔
2.3 物种丰富度和物种密度的垂直梯度
物种丰富度的垂直梯度根据各个海拔段的分段范围以及每种植物分布的海拔范围,利用编程手段,统计每个海拔段内的植物物种及其所属的科、属、种以及捕捉到蝴蝶的数量。
一个地区的物种丰富度受该地区的面积影响较大。为了消除面积的影响,研究地形特征对物种垂直分布的影响时,需要用面积对物种丰富度进行调整。本文使用物种密度(Species Density)这一指标,其计算公式如下[14]:

式中:D为物种密度,R为区域内物种丰富度,A为区域面积。根据各个海拔段的面积以及各个海拔段分布的物种数量,计算各个海拔段的物种密度。
3 结果与分析
3.1 各海拔段地形和物种基本信息
大峪流域各海拔段的地形和物种信息(表1)。

表1 各海拔段地形和物种基本信息
3.2 地形特征沿海拔梯度的分异
从大峪流域的地形模拟图可以看出,大峪流域是一个沿河流形成的沟谷状地形,不同海拔段的面积(投影面积)随海拔的升高逐渐上升(图2)。随海拔的升高,地形复杂度在总体上呈逐渐下降的趋势(图3),其中在海拔1 000~1 200 m与1 400~1 600 m之间地形复杂度有突然降低的又升高的波动。
3.3 物种丰富度和物种密度在海拔梯度上的变化
蝴蝶科的密度基本变化不大,而属、种的密度及总体密度与其丰富度呈相同的变化趋势(图4),即随着海拔的升高,有逐渐降低的趋势,其中中低海拔地区的密度较高,总体密度在低海拔最高,属、种的密度在800~900m之间最高。受地形的影响,在海拔1 300~1 400 m 之间科、属、种及总体密度都有升高的趋势,随之又逐渐降低。
从蝴蝶的科、属、种丰富度以及总物种丰富度沿海拔梯度的变化趋势可以看出(图5),总体上科、属、种及总物种的丰富度沿海拔的升高,呈逐渐降低的趋势,在中低海拔蝴蝶的丰富度最高,其中最大值出现在海拔800~900 m之间,受地形的影响,在海拔1 200~1 400m 之间科、属、种及总体丰富度都有升高的趋势,随之又逐渐降低。

图2 各海拔段面积随海拔的变化
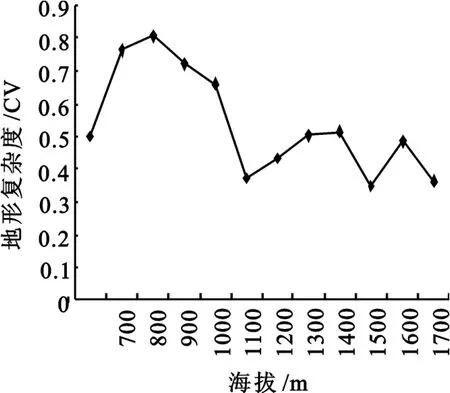
图3 各海拔段地形复杂度随海拔的变化

图4 蝴蝶密度沿海拔梯度的变化

图5 蝴蝶物种丰富度沿海拔梯度的变化
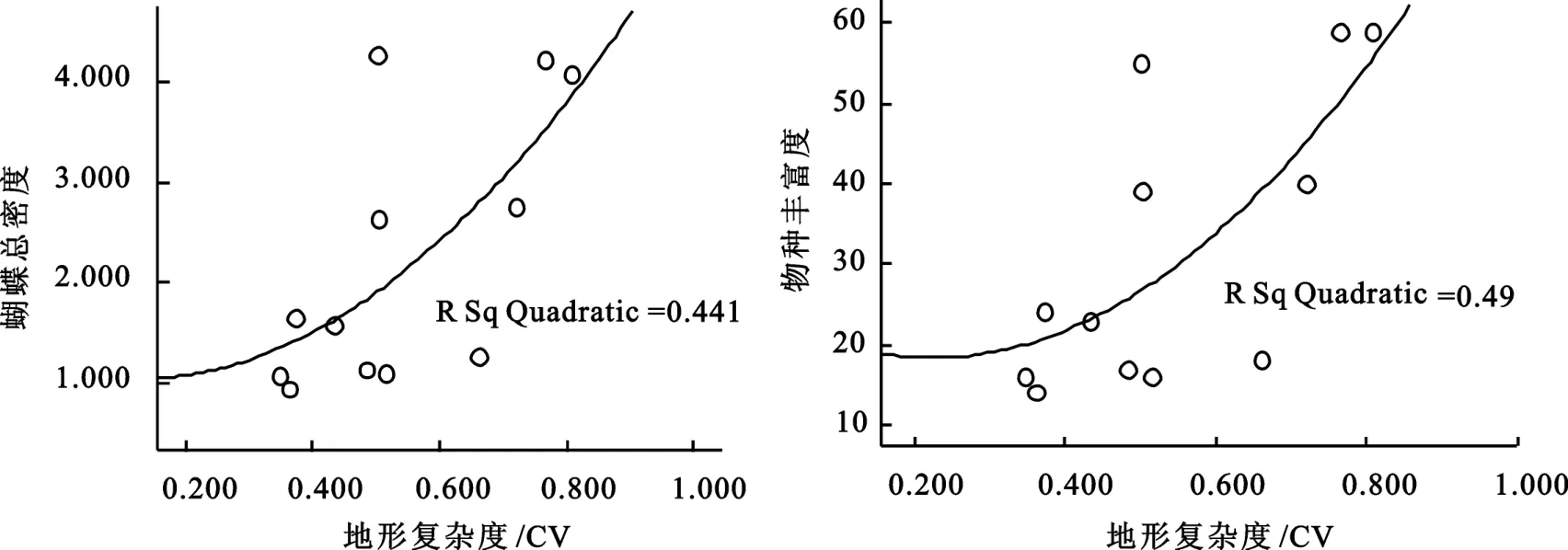
图6 蝴蝶总密度和物种丰富度与地形复杂度散点图
3.4 物种丰富度和物种密度与地形的关系
对蝴蝶总密度和物种丰富度与地形复杂度的数据作散点图(图6),发现散点图大致符合线性回归方程,然后对蝴蝶总密度和物种丰富度与地形作回归分析,其回归方程为:物种丰富度(y=-9.578+76.580· x,R2=0.475,P >0.05),蝴蝶总密度(y=-0.740+5.445·x,R2=0.433,P>0.05)。
从蝴蝶的密度和物种丰富度与地形的关系可以看出,蝴蝶的密度和物种丰富度与地形复杂度呈正相关关系,随着地形复杂度的升高而逐渐升高,但蝴蝶密度和物种丰富度与地形复杂度相关性并不显著。
4 结论与讨论
(1)大峪流域各海拔段的面积随海拔的升高逐渐递增,而地形复杂度总体呈下降的趋势,其中在海拔1 000~1 200 m与1 400~1 600 m之间地形变化较大,面积递增平缓,而地形复杂度有突然降低又升高的波动。
(2)蝴蝶密度和丰富度具有明显的海拔梯度性,随着海拔的升高逐渐降低,其中最大值出现在海拔800~900 m之间,受地形的影响,在海拔1 200~1 400m之间蝴蝶密度和丰富度都有升高的趋势,随之又逐渐降低。
(3)蝴蝶密度和丰富度于地形复杂度呈正相关关系,随地形复杂度的升高而逐渐升高。
物种多样性随海拔梯度变化一直是生态学家感兴趣的论题,海拔梯度包含了温度、水分和光照条件等因素的综合影响[15],一般认为,随着海拔的上升,热量的递减,物种多样性递减。从本文的结果看,大峪流域蝴蝶多样性随海拔升高,蝴蝶多样性下降,但是也应该注意到,其他因子(地形因子)对海拔梯度所框定的水热梯度进行着再修饰,特定海拔高程处生境的海拔梯度效应很可能因为地形等微环境的变化而得到消弱或者加强[16]。
研究表明,地形是影响物种多样性的一个很重要的因子,本文以地形复杂度对蝴蝶多样性进行了分析,总的趋势呈正相关关系,但地形复杂度与蝴蝶多样性相互关系并不显著,其可能原因是在大尺度空间分析中,蝴蝶多样性受气候、生境、寄生植物等因子的影响更大[17],其他因素如系统发育、历史演化、生物地理过程、地质历史等的影响也不可忽略,尤其对于特有种丰富、物种分化剧烈的区域而言[18]。
[1] Paemesan C.Clim ate and species range[J].Natu re,1996,382:765-766.
[2] Erica Leishman,George T.Austin,Andrew D.Weiss An Empirical Test of Rapoport's Ru le:Elevational Gradients in Montane Butterfly Communities[J].Ecplpgy,1998,79(7):2482-2493.
[3] Gen try A H.Changes in Plant Community Diversity and Floristic Composition on Environm ental and Geog raphical Gradients[J].Annals of the Missou ri Botanical Garden,1988,75:1-34.
[4] Theurilat JP,Schlussel A.Vascu lar Plant and Bryophyte Diversity A long Elevation Gradients in the A lps.In:Nagy L,Grabher r G,Koerner C,et al.A lpine Biodiversity in Eu rope[M].New york:Springer-Verlag,2003.
[5] Rahbek C.The elevationalg radient of species richness:a uniform pattern[J].Ecography,1995,18:200-205.
[6] Korner C.Why are thereglobal gradients in species richness Moun tains may hold the answer[J].Trend in Ecology&evolu tion,2000,15:513-514.
[7] W htiaker R H.Classification of natu ral commun ties[M].New York:Amo Press.1977.
[8] Austin M P.Searching for amodel for use in vegetation analysis[J].Vegetato,1980,42:11.
[9] Lieberman D,Lieberman M,Peralta R,et al.T ropical forest structu re and com position on a large-scale altitudinal gradien t in Costa Riea[J].Journal of Ecology,1996,84:137-152.
[10] Lomolino M V.E levational gradients of species-density:historical and prospective view s[J].G lobal Ecology&Biogeogrophy,2001,10:3-13.
[11] B row n JH.M amm als on m ountain sides:elevational patterns of diversity.Global Ecology&Bingengraphy,2001,10:101-109.
[12] 邬伦,刘瑜,张晶等地理信息系统原理、方法和应用[M].北京:科学出版社,2001.
[13] 王志恒,陈安平,方精云.湖南省种子植物物种丰富度于地形的关系[J].地理学报,2004,59(6):889-894.
[14] Qian H.Large-scale biogeographic patterns of vascular plant richness in North America:fillanalysis at the genera level[J].Journal of Biogeography,1998,25:829-836.
[15] 冯建孟,王襄平,方精云.云南独龙江地区种子植物物种多样性垂直分布格局和Rapoport法则的验证[J].北京大学学报,2006,42(4):515-520.
[16] 王国宏.祁连山北坡中段植物群落多样性的垂直分布格局[J].生物多样性,2002,10(1):7-14.
[17] 房丽君,关建玲.蝴蝶对全球气候变化的响应及其研究进展[J].环境昆虫学报,2010,32(3):399-406,311.
[18] 张大才,孙 航.大尺度空间上植物物种丰富度沿海拔梯度分布格局的研究进展[J].西南林学院学报,2009,29(2):74-80.
