川木通与其混伪品和近缘种的ITS2条形码分子鉴定
2011-06-14刘美子李美妮姚辉刘萍
刘美子 李美妮 姚辉 刘萍
川木通为较常用中药,主产于四川等地,具有清热利尿,通经下乳等功效。2010版《中国药典》收载的川木通为毛茛科植物小木通(ClematisarmandiiFranch.)或绣球藤(ClematismontanaBuch.-Ham.)的干燥藤茎[1]。由于本草中记载的异物同名,同物异名和地方用习用品等原因,商品川木通的品种混乱情况较为严重[2],包括毛茛科铁线莲属、木通科木通属(如木通)、马兜铃科马兜铃属(如关木通)等多种植物的藤茎,其中木通与川木通在有效成分、药理作用和主治功能上虽有相似之处[3],但功效各有侧重,应区别使用。关木通由于肾毒性而被禁用后,川木通的需求量日益增大 ,而药典收载的绣球藤生长海拔较高,市场上流通的川木通实际上常混有木通或铁线莲属的多种植物的藤茎[4]。川木通与木通为不同科属的植物,依靠外观形态或显微鉴定可明显区分,但与同属混伪品和近缘种则需要寻找一种快速、准确的分子鉴定方法,以确保临床的安全用药,近年发展起来的DNA条形码技术在近缘种鉴定方面显示出了明显的优势。
DNA条形码技术是随着分子生物学和生物信息学的发展而出现的物种鉴定新技术。它是利用一段短的标准的DNA序列作为标记而建立的物种鉴定方法,目前已被广泛应用于药用植物鉴定[5-8]。nrITS序列常用于不同分类阶元的系统进化研究,尤其是属水平或属下水平的研究[9],nrITS由ITS1、5.8S、ITS2三部分构成,Chase等[10]研究表明,ITS2序列与ITS序列的物种鉴定效率相当,但其序列长度只有ITS的1/3,因此具有较高的扩增效率,ITS2已成为热点的条形码候选序列[11-13]。本文应用ITS2序列对川木通及其混伪品和近缘种进行分析,为近缘物种的分子鉴定提供依据。
1 材料与方法
1.1 材料
川木通基原植物绣球藤及其混伪品木通、三叶木通、棉团铁线莲的的新鲜叶片分别采自四川汶川、成都、南充,广西南宁和北京,由中国医学科学院药用植物研究所林余霖副研究员进行鉴定,硅胶干燥保存,材料信息详见表1。此外,从GenBank下载了川木通与其混伪品和近缘种的ITS2序列,详见表2。

表1 材料采样信息
1.2 方法
采用植物DNA提取试剂盒(Tiangen Biotech Co.,China)提取DNA。ITS2序列的PCR扩增及测定方法均参考Chen等研究[14],将实验样本的双向测序峰图用CodonCode Aligner V3.7.1校对拼接,并剪切引物区。对拼接后得到的序列和GenBank下载的ITS2序列用HMMer注释法切除5.8S和26S端,保留ITS2的全长。用MEGA5软件对川木通种内、种间序列进行分析,采用Koetschan等[15]建立的ITS2数据库预测ITS2二级结构,应用BLAST方法对川木通及其同属混伪品和近缘种进行鉴定分析。
2 结果与分析
2.1 川木通基原植物的ITS2序列长度,GC含量及种内变异
小木通ITS2序列长度为218~220 bp,平均GC含量为68.9%,含5~6个POLY结构(≥5个单碱基重复),为4~5个PolyC和1个PolyG结构。小木通2条序列比对后长度为220 bp,主要变异位点有四处,为74、133位点处的T-C单碱基转换,127位点处的TT-CC双碱基转换和122位点处的C-T单碱基转换,种内K2P距离为0.0285。
绣球藤ITS2序列长度为218 bp,平均GC含量为66.5%,含5个POLY结构,为4个PolyC和1个PolyG。绣球藤3条序列比对后长度为218 bp,主要变异有四处,为23位点处的A-T变异,122位点处的T-C变异,179位点处的ATC-TAA三位点变异,193位点处的A-G变异,种内平均K2P距离为0.0187,种内最大K2P距离为0.0280。
2.2 川木通及其主要混伪品的鉴定
2.2.1 种间变异
川木通与主要同属混伪品女萎(Clematisapiifolia)、短尾铁线莲(Clematisbrevicaudata)的种间平均距离为0.0339,种间最小距离为0.0046, 种间变异较小。 川木通与木通科木通的三个基原木通[Akebiaquinata(Thunb.) Decne.]、三叶木通[Akebiatrifoliata(Thunb.) Koidz.]或白木通[Akebiatrifoliata(Thunb.) Koidz. var. Australis (Diels) Rehd.]的种间平均K2P距离为0.5398,种间最小K2P距离为0.5257,变异位点有94处,种间变异较大;川木通与马兜铃科关木通(AristolochiamanshuriensisKomarov)的种间平均K2P距离为0.6155,种间最小K2P距离为0.5801,变异位点有109处,种间变异较大。川木通与其主要混伪品ITS2序列的种间比对见图1。
2.2.2 ITS2序列二级结构分析
如图2所示,川木通基原植物小木通与绣球藤在螺旋(Helix)IV区有差异,川木通与其同属主要混伪品女萎和短尾铁线莲的差异也在螺旋IV区,而与混伪品木通、三叶木通、白木通在螺旋I、II、III和IV区均存在明显差异。
2.3 川木通与其同属近缘种的鉴定
运用BLAST方法(Chen et al,2010)计算铁线莲属的鉴定效率,结果表明川木通的基原植物小木通和绣球藤均能与同属的31种易混伪品被正确鉴定,而易混品种之间除芹叶铁线莲(C.aethusifolia)与须蕊铁线莲(C.pogonandra),锈毛铁线莲(C.leschenaultiana)与锡金铁线莲(C.siamensis),甘青铁线莲(C.tangutica)与中印铁线莲(C.tibetana)互相不能鉴定外,其他同属近缘物种均能区分开,ITS2条形码对铁线莲属的鉴定成功率达到87%。
3 讨论
川木通的商品药材来源于野生资源,由于生态环境恶化,造成川木通资源短缺[16],于是多种铁线莲属植物的藤茎掺入川木通药材中[4]。大部分文献报道的均是川木通与性状差别较大的木通的鉴定[2,17],而对于亲缘关系较近的同属混伪品,用传统分类方法很难区分,针对此问题,基于ITS2条形码的分子鉴定方法显示出其优势。ITS2作为条形码不仅具备ITS序列变异率高、进化快的特点,还具有保守的5.8S和26S便于设计通用引物,约200~300 bp的长度便于扩增和测序,可以用于部分降解的中药材的鉴定[18]。通过对川木通及其混伪品和近缘物种的分析,川木通基原植物绣球藤和小木通与其混伪品木通和关木通ITS2序列具有显著的种间变异,与同属混伪品女萎和短尾铁线莲的种间变异小。川木通基原植物绣球藤和小木通与同属混伪品女萎和短尾铁线莲的ITS2二级机构在螺旋IV区有差异,用BLAST方法能够将川木通与其同属易混品和近缘物种区分开,对铁线莲属的鉴定成功率达到87%,可见ITS2条形码能够用于亲缘关系较近的正品与混伪品的鉴定。
ITS2序列作为DNA条形码用于中药鉴定具有准确、快捷的优势,此外其二级结构能够为发现新物种和分类学研究提供更多的信息[19],赋予ITS2更为强大的功能,为中药混伪品和近缘物种分子鉴定开拓了新思路。
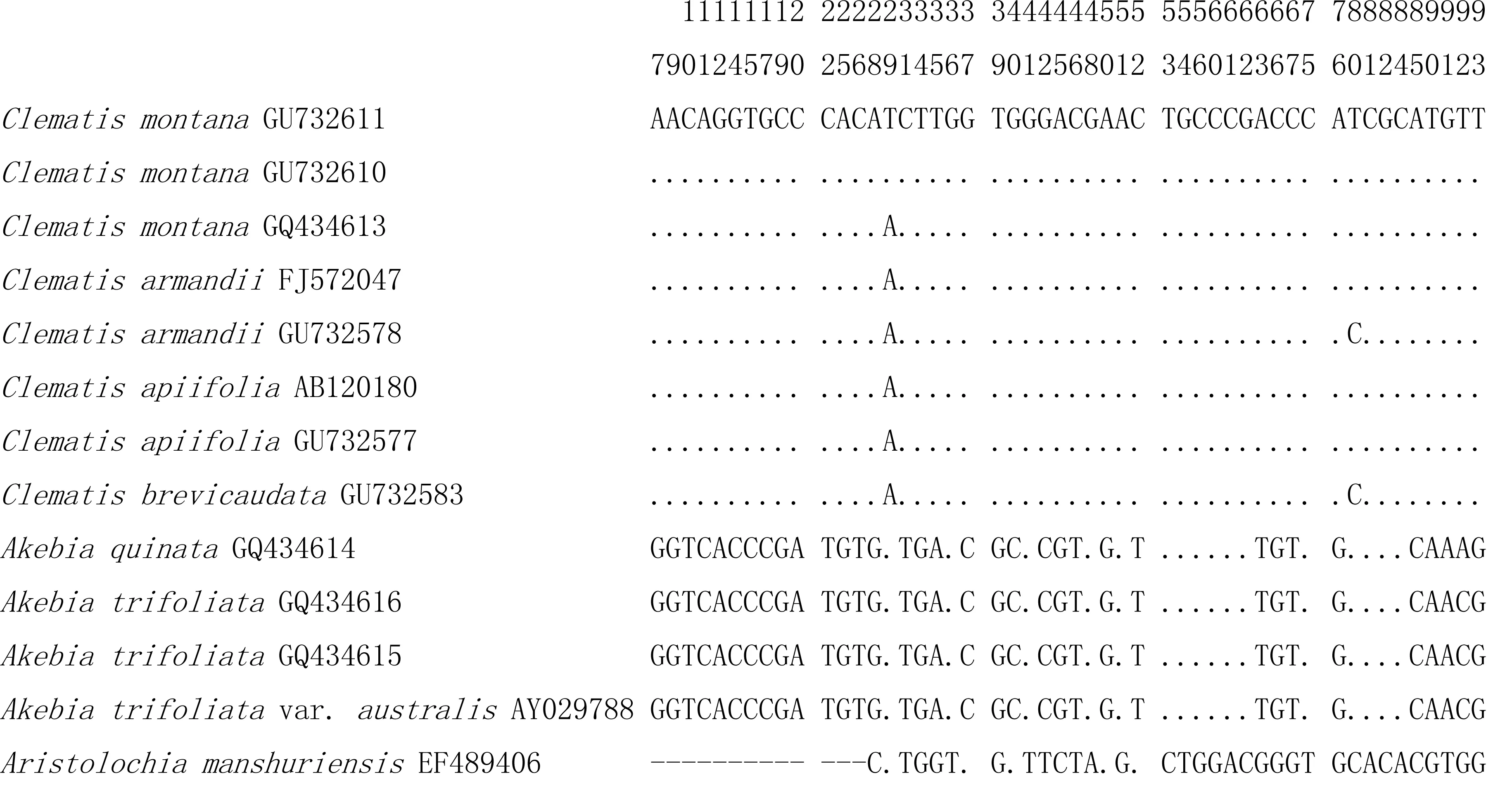


图1 川木通与其主要混伪品木通、女萎、短尾铁线莲的种间变异

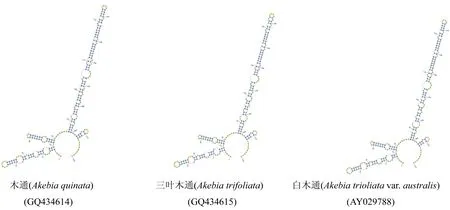
图2 川木通、木通及其主要易混伪品的二级结构比较
[1] 国家药典委员会.中国药典(一部)[M].北京:中国医药科技出版社,2010:34.
[2] 孔增科,李利军,郭明,等.木通与川木通及关木通的鉴别与合理应用[J].河北中医,2007,19(1):58-60.
[3] 何报作.三种被取消药用标准的药材同其替换品及易混淆品的鉴别[J].广西中医学院学报,2005,8(1):48-50.
[4] 唐远,万德光,裴瑾,等.川木通的研究进展[J].时珍国医国药,2007,18(10):2346-2347.
[5] Kress WJ, Wurdack KJ, Zimmer EA,et al. Use of DNA barcodes to identify flowering plants[J]. Proc Natl Acad Sci USA,2005,102(23):8369-8374.
[6] Song JY, Yao H, Li Y,et al. Authentication of the family Polygonaceae in Chinese pharmacopoeia by DNA barcoding technique[J]. J Ethnopharmacol,2009,124(3):434-439.
[7] Gao T, Yao H, Song JY, et al. Identification of medicinal plants in the family Fabaceae using a potential DNA barcode ITS2[J]. J Ethnopharmacol, 2010, 130(1): 116-121.
[8] 陈士林,宋经元,姚辉,等.药用植物DNA条形码鉴定策略及关键技术分析[J].中国天然药物,2009,7(5):322-327.
[9] Alvarez I, Wendel JF. Ribosomal ITS sequences and plant phylogenetic inference[J]. Mol Phylogenet Evol. 2003, 29(3):417-314.
[10] Chase MW,Salamin N, Wilkinson M, et al. Land plants and DNA barcodes: short-term and long-term goals[J]. Philos Trans R Soc Lond B Biol Sci, 2005, 360(1462):1889-1895.
[11] 罗焜,陈士林,陈科力,等.基于芸香科的植物通用DNA条形码研究[J].中国科学:生命科学,2010,30(4):342-351.
[12] 朱英杰,陈士林,宋经元,等.重楼属药用植物DNA 条形码鉴定研究[J].药学学报,2010,45(3):376-382.
[13] Pang XH, Song JY, Chen SL,et al. Applying plant DNA barcodes for Rosaceae species identification[J]. Cladistics, 2011,27:165-170.
[14] Chen S, Yao H, Han JP,et al. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species[J]. PLoS ONE, 2010,5(1):e8613.
[15] Koetschan C, Förster F, Keller A,et al. The ITS2 Database III-sequences and structures for phylogeny[J]. Nucleic Acids Res, 2010,38(Database issue):D275-D279.
[16] 李彬,陈幸方,清茂,等.川木通的品质研究[J].四川中医,2009,27(6):58-59.
[17] 张斌慧,余梅,杨斌.木通、川木通与关木通的比较鉴别[J].贵阳中医学院学报,2006,28(5):37-38.
[18] Chiou SJ, Yen JH, Fang CL,et al.Authentication of medicinal herbs using PCR-amplified ITS2 with specific primers[J].Planta Med, 2007,73(13):1421-1426.
[19] Prasad PK, Tandon V, Biswal DK,et al. Phylogenetic reconstruction using secondary structures and sequence motifs of ITS2 rDNA of Paragonimus westermani (Kerbert, 1878) Braun, 1899(Digenea: Paragonimidae)and related species[J]. BMC Genomics,2009,10(Suppl 3):S25.
