连作对烤烟根际土壤微生物数量和酶活性的动态影响
2011-05-10古战朝习向银刘红杰刘朝科周显波汤书开张重义
古战朝,习向银,刘红杰,刘朝科,周显波,汤书开,张重义,4
(1.河南农业大学,河南郑州450002;2.西南大学,重庆400715;3.川渝中烟工业公司,四川成都610031;4.福建农林大学,福建 福州350002)
烟草是忌连作作物,由于受耕地有限、经济利益驱动和种植条件等因素的制约,烟叶生产耕作制度单一,也即连作或复种连作的面积越来越大,使得连作障碍成为烟草农业生产的突出问题.赵凯等[1]对典型烟区调查发现,烤烟连作3 a以上就会对烟叶的外观与内在质量均产生显著的不良影响.主要表现在中上等烟比例与烟叶的烟碱、还原糖、钾含量以及施木克值、糖碱比和钾氯比等重要化学指标上.晋艳等[2]在1998—2000年对连作烤烟烟叶产量和品质状况的研究结果表明,连作造成烟株株高、叶面积指数、烟叶产量、产值、中上等烟比例、均价等均有不同程度的降低.导致连作障碍的因素相当复杂,如土壤微生物活动、土壤酶活性、土壤根系分泌物[3]、土壤养分状况等.而微生物是土壤物质转化的重要参与者,土壤有机质转化所需能量的90%以上来自微生物的分解作用,是土壤有机质和养分转化与循环的动力[4,5].土壤中微生物的种类较多,有细菌、真菌、放线菌和原生动物等,数量也很大,它们具有较大的比表面积,代谢活动旺盛[6].大部分土壤微生物对作物的生长发育是有益的,它们参与土壤的形成、物质循环和肥力演变,是影响作物生长发育的重要环境条件之一.贾志红等[7]通过提取土壤细菌总DNA,用PCR-DGGE法研究了烤烟轮作和连作对土壤细菌群落多样性的影响,认为连作会导致土壤微生物多样性下降.王茂盛等[8]研究表明,随着烟草连作年限的增加,细菌数量下降,青枯病病原菌数量逐渐减少,真菌数量增加.土壤酶主要来自微生物和植株根系分泌等途径,此外还有土壤动植物活体分泌和动植物残体的分解[9].连作可使土壤中磷酸酶和脲酶活性降低,使蔗糖酶活性增强,而转化酶活性只有连作年限超过5 a后才表现出减少趋势[10].娄翼来等[11]研究发现,随着植烟年限的延长,表层土壤过氧化氢酶、转化酶、脲酶和中性磷酸酶活性的变化均表现为先增强后减弱,而它们在次表层土壤中则均呈现不同程度的降低.于宁等[12]研究表明,与正茬烟田相比,连作烟田的转化酶、脲酶、中性磷酸酶和过氧化氢酶活性显著下降,这将直接影响土壤养分的转化及作物对土壤养分的有效吸收.前人的研究结果表明,烤烟根际土壤受到烤烟根系生长的直接影响,烤烟根际土壤微生物和酶活性的平衡发展是预防烟草连作障碍发生的基础.但之前的研究对生育期内烤烟根际土壤微生物数量和酶活性的动态变化情况研究较少,特别是在受控条件下的研究未见报道.本研究试图通过修建隔离池探讨不同连作年限对烟草移栽后5个时期的根际土壤微生物数量及其酶活性动态变化,为进一步研究烤烟连作对根际土壤微生态系统的影响,探讨烤烟连作障碍的消减技术提供理论依据和技术支撑.
1 材料与方法
1.1 试验设计
该试验于2010年在重庆市武隆县和顺镇青木池村川渝中烟科技示范园隔离池内进行.供试烤烟品种为云烟87.试验设3个处理,分别为:处理1,正茬(未种植过烟草);处理2,连作3 a;处理3,连作8 a.试验用土分别取自正茬、连作3 a和连作8 a的地块耕层土壤.试验地前茬作物一致,均为玉米.试验采用随机区组设计,3次重复,每小区面积5 m ×2.2 m.烤烟移栽期为2010-05-16,行株距为1.2 m×0.55 m,留叶数为21片.肥料种类为烟草专用肥和硝酸钾.m(氮)∶m(磷)∶m(钾)=1∶1.5∶2.5.施纯氮量为 105 kg·hm-2,纯氮 60% 于移栽前条施,40%按当地追肥习惯于团棵期追施.田间管理按重庆市烟草田间管理技术规程执行.试验土壤类型为黄壤,具体养分状况见表1.

表1 不同处理土壤基础养分状况Table 1 Soil nutrient status in different treatments
1.2 根际土样的采集
烤烟根际土壤样品的采集时间分别为移栽后45,65,75,98,113 d.采集方法为抖根法.用 5 点取样法采集土样,混合均匀后用无菌自封袋包装,分2部分保存:一部分保存在4℃冰箱中,供土壤生物数量分析;一部分风干、研磨,过60目筛,供土壤养分及土壤酶活性分析.
1.3 项目分析测定方法
1.3.1土壤酶活性的测定 脲酶采用比色法,其活性用24 h后1 g土壤中NH3-N的毫克数表示;过氧化氢酶采用滴定法,其活性用1 h后1 g土壤的0.1 mol·L-1K2MnO4的毫升数表示;蔗糖酶采用比色法,其活性用24 h后1 g土壤葡萄糖的毫克数表示;磷酸酶采用比色法,其活性用24 h后1 g土壤释放出的酚的毫克数表示[13].
1.3.2 微生物区系分析 细菌在36℃下,在牛肉膏蛋白胨琼脂培养基中培养30 h;放线菌和真菌在28~30℃条件下,分别在高氏1号培养基和马丁氏培养基中培养5 d.
细菌、放线菌和真菌的计数采用平板计数法[14].
1.4 数据分析
采用Microsoft Excel 2007和SPSS 18.0统计软件进行数据统计分析.
2 结果与分析
2.1 连作年限对烟草根际土壤微生物数量的影响
由表2可知,根际土壤中细菌数量在烤烟生育期内呈“升-降-升-降”的变化趋势,且在移栽后65和98 d分别出现1个峰值.不同连作年限处理间相比较,连作3 a的细菌数量最多,且在移栽后98 d前与其它处理间存在极显著性差异,正茬的细菌数量次之.连作8 a的细菌数量最少,且除在移栽后45 d与正茬的无显著性差异外,与其它处理相比,在各个生育时期均存在极显著性差异.各处理根际土壤中真菌数量均在移栽后65 d前逐渐增加到最高峰,之后逐渐下降.在各个生育时期根际土壤中真菌数量均以正茬的最多,最多时达到9.20 ×105个·g-1,连作 3 a的次之,两者在移栽后65和75 d时存在显著性差异,但在各生育时期均与连作8 a的存在显著性差异.各处理间相比较来说,真菌数量随着连作年限的增加呈下降趋势.各处理根际土壤中放线菌数量在烤烟生育期内除连作8 a呈下降趋势外,其余2个处理均先降后升.不同处理间相比,连作3 a的放线菌数量最多,其次为正茬,且除移栽后75 d外,两者放线菌数量在其余各生育时期均与连作8 a的存在极显著性差异.
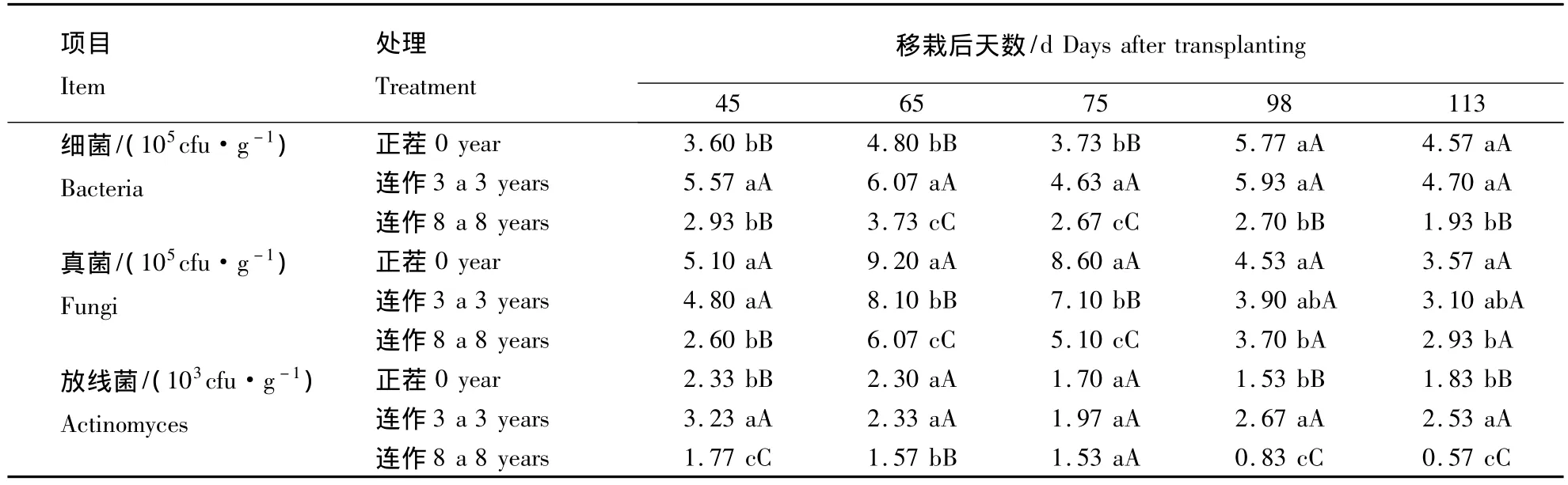
表2 连作年限对烟草根际土壤微生物数量的动态影响Table 2 Effect of microbial quantity in different consecutive cropping years
2.2 连作年限对烟草根际土壤脲酶活性的影响
从图1可以看出,随着烤烟生育期的推进,3个处理的根际土壤脲酶活性大致呈“升-降-升-降”的变化规律,且在移栽后65和98 d分别出现1个峰值.这说明在烤烟生育的旺长期和打顶后,需要较多的营养物质供给,此时脲酶活性显著增强,可以促进尿素的分解,满足烤烟生长的需要.各处理之间,根际土壤脲酶活性在各时期均以连作3 a的最大,正茬次之,连作8 a的最小.在移栽后45 d时,3个处理根际土壤脲酶活性相差较小,但正茬、连作3 a与连作8 a之间存在显著性差异,而正茬与连作3 a之间无显著性差异.随着烤烟的生长发育,正茬和连作8 a的根际土壤脲酶活性的差距又逐渐增大,且差异显著.

图1 连作年限对烟草根际土壤脲酶活性的影响Fig.1 Effect of urease activity in different consecutive cropping years
2.3 连作年限对烟草根际土壤磷酸酶活性的影响
从图2可以看出,连作3 a的根际土壤磷酸酶活性移栽后113 d时,比45 d时下降了7.06%.各处理之间相比,各时期根际土壤磷酸酶活性大小为:连作3 a>正茬>连作8 a.正茬与连作8 a之间除移栽后65 d外,存在显著性差异.在烤烟生育期内,3个处理土壤磷酸酶呈“升-降-升-降”的变化趋势,且在烤烟移栽后65和98 d分别出现1个峰.根际土壤磷酸酶的活性变化与烤烟的生长发育状况相一致,在烤烟生育的旺长期(移栽后65至98 d),需要较多的营养物质供给,此时磷酸酶活性显著增强,可以促进无机磷酸的释放,满足烤烟生长的需要.而在移栽98 d时连作8 a的峰值很小,这可能是由于随着烤烟的生长发育,土壤中有机质含量减少,导致土壤磷酸酶的活性降低[15].

图2 连作年限对烟草根际土壤磷酸酶活性的影响Fig.2 Effect of phosphatase activity in different consecutive cropping years
2.4 连作年限对烟草根际土壤蔗糖酶活性的影响
从图3可以看出,随着烤烟连作年限的增加,根际土壤蔗糖酶活性总体呈曲折下降趋势,其中连作8 a的下降最为明显,移栽后113 d时仅为移栽后45 d时的69.74%.根际土壤蔗糖酶活性与脲酶、磷酸酶的活性变化相似,在烤烟生长的移栽后65和98 d各出现1个峰.各处理之间,根际土壤蔗糖酶活性在烤烟生育各时期均存在显著性差异,其中以连作3 a的最高,正茬的次之,连作8 a的最低.
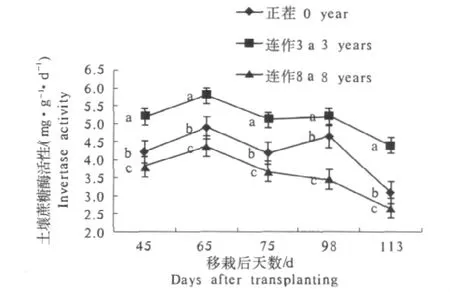
图3 连作年限对烟草根际土壤蔗糖酶活性的影响Fig.3 Effect of invertase activity in different consecutive cropping years
2.5 连作年限对烟草根际土壤过氧化氢酶活性的影响
由图4可见,根际土壤过氧化氢酶活性在烤烟生育期内曲折下降,其中连作3 a的下降最为明显,移栽后113 d时的过氧化氢酶活性仅为移栽后45 d时的63.48%.各处理之间,同一时期土壤过氧化氢酶活性均以连作3 a的最高,正茬的次之,连作8 a的最小,且3个处理之间均存在显著性差异.
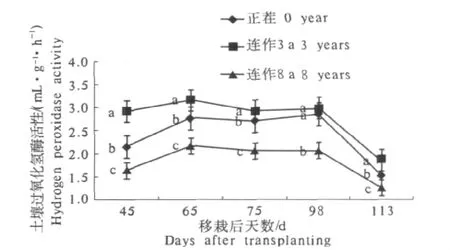
图4 连作年限对烟草根际土壤过氧化氢酶活性的影响Fig.4 Effect of hydrogen peroxidase activity in different consecutive cropping years
3 结论与讨论
微生物是土壤中有机质和养分转化与循环的动力,正常的土壤中微生物数量一般是细菌最多.本试验表明,各处理在整个烤烟生育期内,细菌数量大致呈“升-降-升-降”的变化趋势,放线菌数量除连作8 a呈连续下降趋势外,其余2个处理均先降后升,但均低于移栽初期水平,这与郭红祥等[16]的研究结果一致.不同处理之间比较,各时期根际土壤中细菌对烤烟连作表现敏感,数量下降明显,随着连作年限的增加,真菌数量也有较为明显的下降,放线菌数量先升后降,变化较稳定.李仲强等[17]认为,不同连作年限之间比较,放线菌数量变化之所以较稳定,可能是由于放线菌与细菌相比,生长较慢、受农业技术措施的影响也表现较慢的缘故.总体上根际土壤微生物数量随着连作年限的增加均呈下降趋势,这与胡元森等[18]的研究结果一致,他认为真菌数量的下降可能是由于少数真菌类群富集,多数真菌类群数量减少,种群变化呈单一趋势导致数量减少.本研究表明,不同处理间真菌所占微生物数量的比例,随连作年限的增加有明显的上升趋势,而细菌所占比例有所下降.连作导致根际土壤的细菌数量下降,一方面可能是因为连作导致土壤中养分含量失衡,造成土壤贫瘠,刘训理等[19]的研究表明,土壤肥沃的烟区根际细菌的数量最多,真菌数量较少;土壤贫瘠烟区根际真菌的数量较多,细菌数量较少;另一方面可能是由于土壤根系分泌物浓度抑制土壤微生物生长发育造成的,而细菌和放线菌对此反应较为敏感,真菌对此反应不太敏感,导致土壤微生物中真菌比例增加.由此可以看出,连作虽降低了土壤细菌、真菌等微生物的数量,但由于细菌数量降低明显,导致烤烟根际微生物中真菌的比例反而升高,使土壤微生物区系从高肥的“细菌型”向低肥的“真菌型”转化,这与于贵瑞[20]、于广武[21]和邹莉等[22]研究连作大豆土壤微生物种群变化时得出的结论一致.
连作可能导致土壤中某一种养分过度消耗,结果迫使这些元素对作物的供应越来越不足,造成土壤养分的过度消耗[23].作物主要生育时期内,不同耕作方式下的土壤酶活性变化趋势与作物的生长及需肥规律基本一致[24],土壤酶活性能够体现土壤的生物学活性,表征土壤的综合肥力及土壤养分转化进程[25],人们通常用脲酶、磷酸酶、蔗糖酶、过氧化氢酶等酶活性来评价土壤肥力和肥效.在本研究中,土壤脲酶、磷酸酶、蔗糖酶活性以连作3 a最高,其次为正茬,连作8 a最低,且三者存在显著性差异,结果表明,随着连作年限的增加,土壤酶活性先升高后下降.这说明,短期连作使土壤酶活性升高,而长期连作会降低土壤酶活性.这与根际土壤微生物数量变化相一致,说明土壤酶活性随土壤微生物数量和有机质含量的变化而变化.在烤烟生育期内,随着烤烟生育期的推进,土壤酶活性大致呈“M”型的变化趋势,且在移栽后65和98 d分别出现1个峰值,这亦可能是由于在烤烟的旺盛生长期内,需要吸收大量的营养物质,土壤酶活性的变化规律与烤烟生长特性表现出了一定的协调性.
薛小平等[26]研究认为,短期连作的烤烟根际土壤微生物群落与原始土壤的相比较,更适宜于烟株的生长.李坤等[27]的研究表明,葡萄短期种植根际土壤细菌和真菌的多样性增加,且根围土壤细菌和真菌的种群结构不同于空地土壤.烤烟的根系分泌物为其次生代谢产物(化感物质),张重义等[28]认为化感物质引起植物产生自毒作用,是导致烤烟连作障碍的主导因子,自毒作用的方式具有浓度效应的特征.刘新虎等[29]的研究也发现,低浓度的根系分泌物对棉花幼苗植株的根长、株高、地下部分鲜重、地上部分鲜重有明显的促进作用,而高浓度则有抑制作用.根系分泌物作为根际微生物的营养来源之一,密切影响其种类和数量.由此推测,烤烟短期连作(小于3 a)情况下烤烟生长与土壤根际微生物群落关系密切,而长期连作与化感物质引起的连作障碍效应关系密切,该推测还有待进一步研究.
[1] 赵 凯,娄翼来,王玲莉,等.烤烟连作对烟叶产量和质量的影响[J].现代农业科技,2008(8):118-119.
[2] 晋 艳,杨字虹,段玉琪,等.烤烟连作对烟叶产量和质量的影响研究初报[J].烟草科技,2002(1):41 -45.
[3] 陈冬梅,柯文辉,陈兰兰,等.连作对白肋烟根际土壤细菌群落多样性的影响[J].应用生态学报,2010,21(7):1751-1758.
[4] 李阜棣.土壤微生物学[M].北京:中国农业出版社,1995.
[5] MOONY J H,CUSHMAN E,MEDINA O H,et al.Functional role of biodiversity[M].New York:John Wiley& Sons Ltd,1996.
[6] WHITING S N,SOUZA D M,TERR Y N.Rhizosphere bacteria mobilize Zn for hyperaccumulation by Thlaspi caerulescens[J].Science and Technology,2001,35:3144-3150.
[7] 贾志红,易建华,苏以荣,等.烟区轮作与连作土壤细菌群落多样性比较[J].生态环境学报,2010,19(7):1578-1585.
[8] 王茂盛,姜超英,潘文杰,等.不同连作年限的植烟土壤理化性质与微生物群落动态研究[J].安徽农业学报,2008,36(12):5033-5034:5062.
[9] 张咏梅,周国逸,吴 宁.土壤酶学的研究进展[J].热带亚热带植物学报,2004,12(1):83-90.
[10]彭有才,刘 挺,赵俊杰,等.连作对土壤性状影响的研究进展[J].江西农业学报,2009,21(9):100 -103.
[11]娄翼来,关连珠,王玲莉,等.不同植烟年限土壤pH和酶活性的变化[J].植物营养与肥料学报,2007,13(3):531-534.
[12]于 宁,娄翼来,严 丽,等.施石灰对北方连作烟田土壤酸度调节及酶活性恢复研究[J].土壤通报,2008,39(4):849-851.
[13]关松荫.土壤酶学研究方法[M].北京:农业出版社.1986.
[14]中国科学院南京土壤研究所微生物室.土壤微生物研究法[M].北京:科学出版社,1985.
[15]程 伟,隋跃字,焦晓光,等.土壤有机质含量与磷酸酶活性关系研究[J].农业系统科学与综合研究,2008,24(3):305 -307.
[16]郭红祥,刘卫群,姜占省.施用饼肥对烤烟根系土壤微生物的影响[J].河南农业大学学报,2002,36(4):344-347.
[17]李仲强,谭周进,夏海鳌.耕作制度对土壤微生物区系的影响[J].湖南农业科学,2001(2):24-25.
[18]胡元森,刘亚峰,吴 坤,等.黄瓜连作土壤微生物区系变化研究[J].土壤通报,2006,37(1):126-129.
[19]刘训理,王 超,吴 凡,等.烟草根际微生物研究[J].生态学报,2006,26(2):552 -557.
[20]于贵瑞,陆欣来,韩静淑,等.大豆、向日葵等作物连作障碍与轮作效应机理研究初探[J].生态学杂志,1988,7(2):1 -8.
[21]于广武,许艳丽,刘晓冰,等.大豆连作障碍机制研究初报[J].大豆科学,1993,12(3):237 -243.
[22]邹 莉,袁晓颖,李 玲,等.连作对大豆根部土壤微生物的影响研究[J].微生物学杂志,2005,25(2):27 -30.
[23]刘晓冰,于广武,许艳丽,等.大豆连作效应分析[J].农业系统科学与综合研究,1990,3(3):40-44.
[24]姜桂英,黄绍敏,郭斗斗.不同耕作和轮作方式下作物生育期内土壤酶活性的动态变化特征[J].河南农业大学学报,2009,43(3):335 -342.
[25]陈宏宇,李晓鸣,王敬国.抗病性不同大豆品种根面及根际微生物区系的变化Ⅱ.连作大豆(重茬)根面及根际微生物区系的变化[J].植物营养与肥科学报,2006,12(1):104 -108.
[26]薛小平,张 琴,陈 懿,等.施肥和微生物对连作烟苗生长的影响[J].江西农业学报,2008,20(9):1 -4.
[27]李 坤,郭修武,孙英妮,等.葡萄连作对土壤细菌和真菌种群的影响[J].应用生态学报,2009,20(12):3109-3114.
[28]张重义,谢小波,王 毅,等.烟草化感自毒作用与其连作障碍研究的启示[J].中国烟草学报,2011,17(4):8-13.
[29]刘新虎,赵小亮,万传星,等.棉花根系分泌物对棉苗生长及生理活性的影响[J].棉花学报,2009,21(4):335-336.
