白花树不同种源苗期光响应特性研究
2011-05-09李因刚柳新红朱光权
赵 勋,李因刚,柳新红*,朱光权,徐 梁
(1. 浙江农林大学,浙江 临安 311300;2. 浙江省林业科学研究院,浙江 杭州 310023)
白花树不同种源苗期光响应特性研究
赵 勋1,2,李因刚2,柳新红2*,朱光权2,徐 梁2
(1. 浙江农林大学,浙江 临安 311300;2. 浙江省林业科学研究院,浙江 杭州 310023)
以贵州从江、江西大余、湖南宜章、福建南平、广西上思、云南屏边6个白花树种源的1年生苗为研究对象,利用LI-6400便携式光合作用测定系统对其光合特性进行研究。结果表明,白花树6个种源的生长高度增长量存在明显差异;云南屏边种源的光饱和点(LSP)、最大净光合速率(Amax)、表观量子效率(AQY)均高于其它种源,而暗呼吸速率(Rd)低于其他种源;白花树6个种源净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)及水分利用效率(WUE)对不同光合有效量子通量密度(PPFD)的响应均有一定程度的差异。通径分析表明,影响白花树不同种源苗期叶片净光合速率的主要因子不完全相同,但胞间CO2浓度却是各个种源共有的主要影响因子。
白花树;种源;生长量;光合—光响应
光合作用是植物生长发育形成的基础,也是决定植物长势优劣的主要因素之一,同时又受内外因素的影响,而光照强度是影响植物光合作用的重要外界环境因素。植物的光合性能可以通过光合速率、蒸腾速率、气孔导度、水分利用效率和胞间CO2浓度等得到衡量。研究植物的光合特性,有利于了解植物对光能的利用效率,阐明植物光合的生态学特征。宋宏伟等[1]对不同种源黄连木的光合特性进行研究,选择出了苗期表现优良的种源;杨全等[2]以8个种源黄芩为研究对象,对其光合特性进行了比较研究,结果表明:8个种源黄芩的净光合速率、光饱和点、光补偿点等光合指标间差异显著;何小勇等[3]对3种紫金牛属植物光合光响应特性的研究,表明2种植物适应强光的能力、生长势等显著不同。
白花树(Styrax tonkinensis)属安息香科安息香属落叶小乔木,为热带、亚热带树种[4]。以往对该树种的研究主要集中在工业用材、药用与生物质能源开发和野外调查等方面[5~9],近年李因刚等[10]在白花树人工林与优树选择方面进行研究,而对白花树的光合特性方面研究尚未见报道。通过对白花树光合特性的研究,揭示其生理学特性,探讨光合特性的种源变异,将有助于探明白花树不同种源的光合能力以及对光照强度的生理响应,进而为白花树的栽培和良种选育提供理论与实践指导意义。
1 材料与方法
1.1 试验地概况
试验地位于杭州市西湖区浙江林业科学研究院试验苗圃内,属亚热带季风性气候,四季分明,温暖湿润。年平均气温15.9~17.0℃,呈南高北低分布。极端最高气温39.8~42.9℃,极端最低气温-7.1 ~-15.0℃。年平均相对湿度76%~81%。无霜期199~328 d。土壤类型为山地红壤,砾石含量较高,土壤较贫瘠,pH6.0。
1.2 试验材料
观察测定的白花树于2010年3月播种育苗,行距20 cm,株距20 cm。苗木出土后,施肥1次,除草3次,管理一般。
1.3 测定方法
1.3.1 光合—光响应曲线测定 试验于2010年9月进行,选择晴天,于9:30-11:30用LI-6400便携式光合作用测定系统(LI-COR, Lincoln, Nebraska, USA)的自动光曲线程序来测定光合—光响应,每次选取正常生长植株,向阳中部枝条的第5、第6片生长正常的叶片;每个种源选取3株,每株测定3个叶片。光源为LI-6400红蓝LED光源,控制LI-6400参比室的CO2浓度为400 μmol·mol-1,在光合有效量子通量密度(PPFD)分别为2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、150、100、50、20、0 μmol·m-2·s-1的条件下测定白花树6个种源成熟叶片的净光合速率(Pn, μm o l·m-2·s-1CO2)、蒸腾速率(T r,mmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、胞间CO2浓度(Ci,μmol·mol-1)、叶片温度(Tleaf,℃)等光合生理参数。分析计算光饱和点(LSP)、光补偿点(LCP)、最大净光合速率(Amax)、暗呼吸速率(Rd)、表观量子效率(AQY)、叶片瞬时水分利用效率(WUE)等参数。
1.3.2 苗高生长量测定 分别于2010年9月1-30日,每隔10 d,测定白花树6个种源生长高度。每个重复选择10株,共3个重复。每株挂上标签标记,每次测相同植株。
1.4 数据统计分析方法
应用SPSS16.0和SAS8.1统计分析软件进行数据分析处理,应用Excel 2003软件进行图形和表格处理。
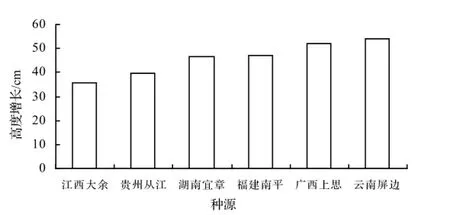
图1 白花树不同种源9月苗高生长量Figure 1 Height grow th ofS. tonkinensisfrom different provenances in Sep.
2 结果与分析
2.1 白花树不同种源的苗高生长量
植物生长高度是植物生长状况的主要的表征因子之一,也是植物生长快慢的主要表现形式之一。由图1可见知,9月白花树苗木的高生长平均达到了39.1 cm,生长迅速。虽然白花树的苗高生长在种源间没有显著差异,但是种源间差异比较明显,其中生长量最大的云南屏边种源达到了48.3 cm,最小的江西大余种源只有32.3 cm,两者相差0.50倍。6个种源9月的苗高生长量大小顺序是:云南屏边>广西上思>福建南平>贵州从江>湖南宜章>江西大余。
2.2 白花树不同种源的光合—光响应参数
光合能力的强弱在一定程度上取决于物种的遗传特性。观测结果表明,不同种源白花树的光合光响应特性存在一定的差异,结果见表1。

表1 白花树不同种源光响应参数Table 1 Light response parameter among different provenance of S. tonkinensis
LSP反映了植物对强光的喜好程度,LSP值越高说明植物在受到强光刺激时越不易发生光抑制[11]。从表1中可以看出,不同种源的LSP存在显著差异。白花树云南屏边和广西上思种源的LSP显著高于江西大余种源,这表明原产地在云南屏边、广西上思的白花树对强光的利用能力高于江西大余种源。这与9月云南屏边、广西上思种源苗木的生长明显大于江西大余种源是相一致的。
LCP是植物利用弱光能力的一项重要指标,LCP越小表明对弱光的利用能力相对较强[12]。6个种源中,湖南宜章种源与贵州从江、江西大余种源的LCP有显著差异。这表明湖南宜章种源对弱光利用能力强于贵州从江和江西大余种源,而湖南宜章种源与福建南平、广西上思和云南屏边种源之间,在弱光利用能力方面没有显著差异。同样可以发现,贵州从江、江西大余、福建南平、广西上思、云南屏边等地种源,在弱光利用能力上没有显著差异。6个种源的LCP都没有极显著差异,这与9月生长高度增加量上,没有显著差异相一致。
从光照利用范围来看,云南屏边种源利用范围最大为578.855μmol·m-2·s-1,依次是广西上思种源570.320 μmol·m-2·s-1、福建南平种源560.807μmol·m-2·s-1、贵州从江种源527.938μmol·m-2·s-1、湖南宜章种源521.016μmol·m-2·s-1、江西大余种源483.492μmol·m-2·s-1。
Amax反映叶片的光合潜能。从表1可看出,云南屏边种源光合速率最大,江西大余种源最低,且与福建南平、广西上思、云南屏边种源有显著差异。光照利用范围大,有利于光合产物的积累,越有利于植物的生长。这与白花树云南屏边种源苗期高度增长量高于其他种源相符合。
AQY反映植物对弱光的利用效率[13],AQY值越高,对弱光的利用效率就越高。白花树的6个种源中,云南屏边种源AQY与其他种源有极显著差异。从AQY上也可说明,9月份云南屏边种源在高度增长量上,明显高于其余5个参试种源。
暗呼吸速率Rd反映了无光照时呼吸消耗。一般情况下,暗呼吸速率高,不利于植物的生长。同样可知,云南屏边种源与贵州从江种源、江西大余种源以及福建南平种源有极显著性差异,湖南宜章种源与贵州从江种源、江西大余种源存在显著性差异。6个种源中,云南屏边种源Rd最低。这从暗呼吸速率方面证实,白花树云南屏边种源9月生长高度增长量,高于其他5个参试种源。
2.3 白花树不同种源的光合参数对光合有效辐射的响应
2.3.1 净光合速率对光合有效辐射的响应 植物叶片的光合作用是植物物质生产的基础,光合速率的高低决定了光合能力的强弱,是植物生物产量形成的关键。从光合—光响应曲线(图2)可知,白花树不同种源初始阶段的光合速率在0~500μmol·m-2·s-1强度的范围内均随光照强度的增强而增加,并表现出线性相关,且幅度较大而后逐渐变缓,达到一定数值(即光饱和点)后,净光合速率变化较小,都维持在较高水平。在不同的光照强度下,云南屏边的白花树光合能力均表现为最强。
2.3.2 水分利用效率对光合有效辐射的响应 水分利用效率是指植物蒸腾消耗单位质量水分所同化CO2的量。其决定于植物光合与蒸腾速率作用的比例[14],也是影响植物光合作用的主要生理因子之一。由图3可知,白花树不同种源的WUE随PPFD的增大呈抛物线状变化。在起始阶段WUE都随PPFD强度的增加逐渐增大,当达到一定光合有效辐射(光饱和点)以后,变化平缓。起始阶段云南屏边种源WUE最大,其次依次是湖南宜章种源、贵州从江种源、福建南平种源、江西大余种源以及广西上思种源。光饱和点后各种源WUE有所变化,除湖南宜章种源升高为最高和江西大余种源降为最低外,其他种源WUE随PPFD变化与初始阶段相同。

图2 净光合速率对光合有效辐射的响应曲线Figure 2 Photosynthetic response curves of Pn to photosynthetic available radiation

图3 水分利用效率对光合有效辐射的响应曲线Figure 3 Response curves of WUE to photosynthetic available radiation
2.3.3 气孔导度对光合有效辐射的响应 气孔是叶片与外界进行气体交换的门户,气孔的开闭程度对气体交换会产生显著的影响。气孔导度与蒸腾速率成正比,与气孔阻力成反比。由图4可见,6个种源的气孔导度随光合有效辐射的变化趋势基本一致,但广西上思种源与其他种源在绝对量上有明显差异,云南屏边、福建南平、贵州从江、江西大余种源的气孔导度相当,湖南宜章种源最低。
2.3.4 胞间CO2浓度对光合有效辐射的响应 Ci是外界CO2气体进入叶肉细胞过程中所受各种驱动力和阻力,以及叶片内部光合作用和呼吸作用的最终平衡结果。驱动力主要由叶片内外的CO2浓度差来体现[3]。CO2是植物进行光合作用必须的物质之一,Ci低,则不利于光合作用的正常进行,光合速率下降;反之,光合速率提高的潜力大。由图5可知,光能利用Ci的变化趋势是开始时随PPFD的增大而减小,而后下降缓慢逐渐趋于平衡。6个不同种源白花树的Ci变化相似,没有明显差异。

图4 气孔导度对光合有效辐射的响应曲线Figure 4 Response curves of Gs to photosynthetic available
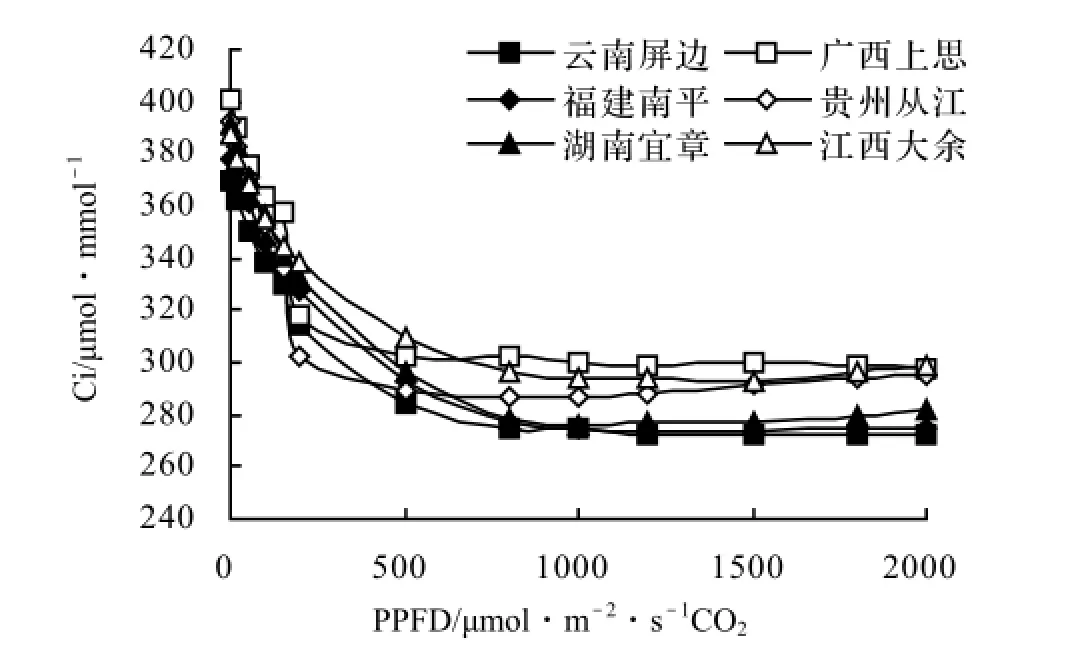
图5 胞间CO2浓度对光合有效辐射的响应曲线Figure 5 Response curves of Ci to photosynthetic available radiation
2.3.5 蒸腾速率对光合有效辐射的响应 蒸腾作用是水分从活的植物体表面(主要是叶片),以水蒸汽状态散失到大气中的过程。与物理学的蒸发过程不同,蒸腾作用不仅受外界环境条件的影响,而且还受植物本身的调节和控制,因此它是一种复杂的生理过程。由图6可见,随着瞬时PDFF的增加,白花树不同种源的Tr均随之增加。广西上思种源蒸腾作用最强,湖南宜章种源蒸腾作用最弱。其他种源蒸腾作用相近。
2.3.6 叶温下蒸汽压亏缺对光合有效辐射的响应 由图7可见,白花树不同种源的叶温下蒸汽压亏缺的变化趋势相似,变化平缓,都集中在1.48~2.0 kPa。白花树湖南宜章种源的叶温下蒸汽压亏缺最低。江西大余种源起始阶段低于广西上思种源,光饱和点后,逐渐高于其他种源。

图6 蒸腾速率对光合有效辐射的响应曲线Figure 6 Response curves of Tr to photosynthetic available radiation
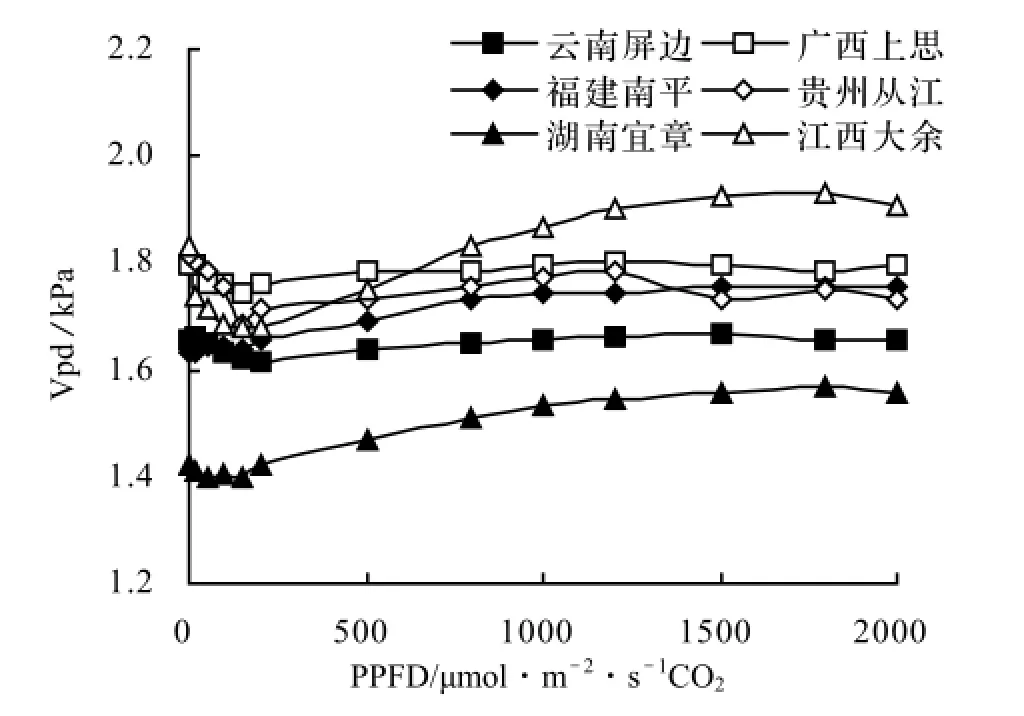
图7 叶温下蒸汽压亏缺对光合有效辐射的响应曲线Figure 7 Response curves of Vpd to photosynthetic available radiation
2.4 影响不同种源净光合速率的生理因子分析 通径分析是研究自变量之间、自变量与因变量之间相互影响关系,描述各个自变量的直接和间接影响程度[15]。为进一步了解以上各光合因子对净光合速率的影响,选择了气孔导度(Gs, 设为X1)、胞间CO2浓度(Ci,设为X2)、蒸腾速率(Tr,设为X3)、叶温下蒸汽压亏缺(Vpd,设为X4)、光合有效辐射(PPFD,设为X5)5个因子为自变量,净光合速率(Pn,设为Y)为因变量,利用逐步通径分析的原理进行相关分析,分析结果见表2。
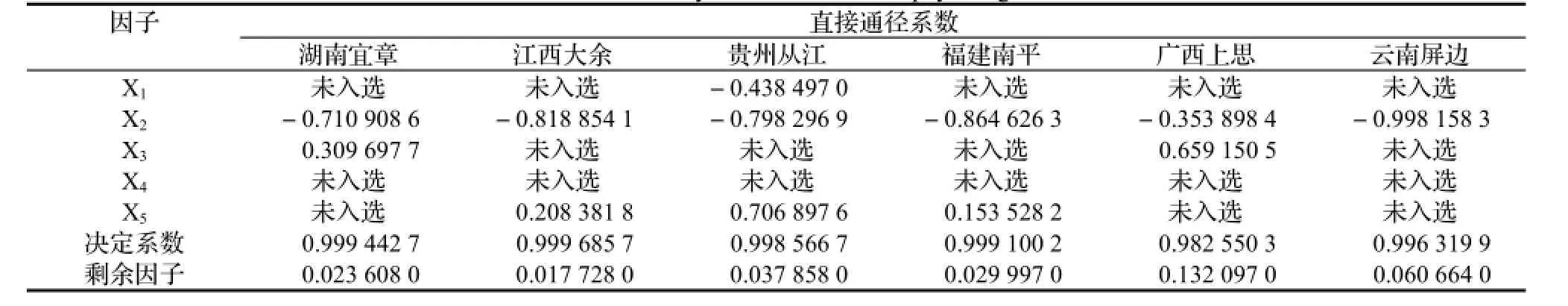
表2 净光合速率与生理因子相关分析Table 2 Correlation analysis between Pn w ith physiological factors
根据表2通径分析结果,显著影响白花树湖南宜章种源叶片净光合速率的因子是胞间CO2浓度和蒸腾速率,而且胞间CO2浓度影响程度高于蒸腾速率;显著影响白花树江西大余种源叶片净光合速率的因子是胞间CO2浓度和光合有效辐射,且胞间CO2浓度影响程度高于净光合速率;显著影响白花树贵州从江种源叶片净光合速率的因子是气孔导度、胞间CO2浓度和光合有效辐射,影响顺序是胞间CO2浓度>净光合速率>气孔导度;显著影响白花树福建南平种源叶片净光合速率的因子是:胞间CO2浓度和光合有效辐射,且胞间CO2浓度影响程度高于净光合速率;显著影响白花树广西上思种源叶片净光合速率的因子是蒸腾速率和胞间CO2浓度,且蒸腾速率影响程度高于胞间CO2浓度;显著影响白花树云南屏边种源叶片净光合速率的因子只有胞间CO2浓度。从分析结果还可发现,胞间CO2浓度对净光合速率的影响,在所有参试种源中,都被入选。陈根云等[16]研究认为,Ci的大小取决于4个可能变化的因素:叶片周围空气的CO2浓度、气孔导度、叶肉导度(gm)和叶肉细胞的光合活性,Ci与Pn可以是正相关,也可以是负相关或无相关。通过本次试验可知,Ci与Pn是负相关,Ci降低是叶肉细胞光合活性增大乃至光合速率增高的结果。
3 结论
(1)同一生境中白花树不同种源的Pn、Tr、Ci、Gs、WUE对PPFD的响应均有不同程度的差异。
(2)云南屏边种源的LCP、Amax、AQY均高于其他种源,表明其光合利用综合能力强于其他种源,其次是广西上思种源,其他依次是福建南平、贵州从江、湖南宜章和江西大余种源。
(3)各种源的光合生理参数的变化与生长高度增长量变化相符合。
(4)6个种源白花树虽属于同种植物,但不同种源间光合生理特性却存在不同程度的差异。这可能是由于自然选择使得各种源已适应原产地生长环境而形成各自的生理特性的结果。引种选育时要选择Pn、LSP、AQY较高,而LCP和Rd较低的种源,这将会更大效率的利用自然光照条件。
(5)蒋高明等[17]认为,植物光合速率除了种间的差异外,还表现在生态型的差异上。通过本试验表明:不同种源白花树的确存在这种差异。影响白花树不同种源苗期叶片净光合速率的主要因子不完全相同,然而胞间CO2浓度却是各个种源共有的主要影响因子。
4 讨论
LSP和LCP是植物的2个重要的光合生理指标,一般LCP和LSP均较低是典型的耐荫植物,能充分地利用弱光进行光合作用;LCP较低而LSP较高的植物对环境适应性很强;LCP较高而LSP较低的植物,对光照的适应范围相对较窄;LCP和LSP都较高的是典型阳性植物[18]。6个不同种源的白花树LCP和LSP总体变化范围是:18.773~ 601.717μmol·m-2·s-1,与杨树、悬铃木、喜树等树种相比,LSP、LCP相对偏低。而有资料记载白花树为阳性树种,这可能是由于本次试验测定有苗木全部是从低纬度的原产地引种到高纬度的浙江杭州播种育苗引起的,另一原因试验材料处于幼苗期,对光的利用能力还没有达到最大水平。
Farquhar等[19~20]认为,Ci的大小是评判气孔限制和非气孔限制的依据之一,Pn、Gs和Ci同时下降时,Pn的下降为气孔限制。相反,如果叶片Pn降低而Ci升高,说明光合作用的限制因素可能是非气孔限制。本试验中6个不同种源的白花树拟合出的Pn预期值,表现出的是在高光合有效辐射照射后的Pn值变化幅度比较平缓,这种现象究竟是气孔限制还是非气孔限制,仍需要进一步的研究;相似的变化趋势还表现在Pn、Tr、Ci、Gs、WUE,而大多数物种是在高光合有效辐射照射时,表现出光合生理指标的下降,可能正是这个原因才使得该树种成为速生树种。以上可能性有待于今后做深一步研究。
[1] 宋宏伟,于海燕,卢绍辉,等. 不同种源黄连木苗期光合特性的研究[J]. 林业科学研究,2009,22(3):454-457.
[2] 杨全,王文全,张卉,等. 8个种源黄芩光合特性的比较研究[J]. 吉林农业大学学报,2006,28(5):530-533.
[3] 何小勇,练发良,李因刚,等. 3种紫金牛属植物光合光响应特性的研究[J]. 浙江林业科技,2008,28(1):15-18.
[4] 中国科学院中国植物志编辑委员会.中国植物志第六十卷(第二分册)[M]. 北京:科学出版社,2004. 84-86.
[5] 柳新红,李因刚,何小勇. 白花树研究进展[J]. 浙江林业科技,2008,28(5):61-65.
[6] 李隆云,肖小河,秦松云,等. 道地药材的形成与分化探讨[J]. 中国中医药科技,1999,6(2):104-106.
[7] 文彬. 西双版纳热带能源植物资源的研究[J]. 国土与自然资源研究,2007(2):84-86.
[8] 孟中贵,周正. 越南安息香引种栽培技术[J]. 中草药,1992,15(4):7-8.
[9] 吴克选,曾志光,周小平,等. 东京野茉莉野外调查报告[J]. 江西林业科技,2002(2):25-27.
[10] 李因刚,周小平,柳新红,等. 越南安息香人工林生长与优树选择的初步研究[J]. 浙江林业科技,2010,30(3):24-28.
[11] 赵溪竹,姜海凤,毛子军. 长白落叶松、日本落叶松和兴安落叶松幼苗光合作用特性比较研究[J]. 植物研究,2007,27(3):361-366.
[12] 巨关升,武菊英. 观赏狼尾草光合特性的研究[J]. 核农学报,2005,19(6):451-455.
[13] 张津林,张志强,查同刚,等. 沙地杨树人工林生理特性[J]. 生态学报,2006,26(5):1 521-1 532.
[14] 杜菁昀,杜占池,崔骁勇. 内蒙古典型草原地区常见植物光合、蒸腾速率和水分利用效率的比较研究[J].草业科学,2003,20(6):11-15.
[15] 杜家菊,陈志伟. 使用SPSS线性回归实现通径分析的方法[J]. 生物学通报,2010,45(2):4-6.
[16] 陈根云,陈娟,许大全. 关于净光合速率和胞间CO2浓度关系的思考[J]. 植物生理学通讯,2010,46(1):64-66.
[17] 蒋高明. 植物生理学[M]. 北京:高等教育出版社,2004. 59, 65.
[18] 冷平生,杨晓红,胡悦,等. 5种园林树木的光合和蒸腾特性的研究[J]. 北京农学院学报,2000,15(4):13-18.
[19] Farquhar G D,Sharkeyt D. Stomatal conductance and photosynthesis[J]. Ann Rev Plant Physiol,1982(33):317-345.
[20] 柯世省. 桂花光合特性的光温响应[J]. 生命科学研究,2007,11(2):110-115.
Study on Light Response Characteristics o f Styrax tonkinensis Seed ling from Six Provenances
ZHAO Xun1,2,LI Yin-gang2,LIU Xin-hong2*,ZHU Guang-quan2,XU Liang2
(1. Zhejiang A & F University, Lin’an 311300, China; 2. Zhejiang Forestry Academy, Hangzhou 310023, China)
Studies were conducted on light response characteristics of 1-year Styrax tonkinensis seedling form six different provenances (Congjiang of Guizhou, Dayu of Jiangxi, Yizhang of Hunan, Nanping of Fujiang, Shangsi of Guangxi, Pingbian of Yunnan) w ith Li-6400 infrared gas analyzer. The results showed that height increment of seedling from six provenances was evident different. Light saturation point (LSP), the maximum net photosynthetic rate (Amax) and apparent quantum yield (AQY) of provenance from Pingbian, Yunnan province was higher than that from the other provenances, only dark respiration rate (Rd) was lower than the others. The net photosynthetic rate (Pn), intercellular CO2concentration (Ci), stomatal conductance (Gs), transpiration rate (Tr) and water use efficiency (WUE) of the six provenances showed different responses to the photosynthetic available photo flux density (PPFD). Path Analysis showed that major factors influencing seedling Pn from different provenances were different, but Ci was the same influencing factor in all provenances.
Styrax tonkinensis; provenance; increment; photosynthesis and light response
S718.51
A
1001-3776(2011)03-0001-06
2010-09-11;
2010-11-29
浙江省重大科技专项“食用油料与能源植物定向培育及开发利用研究”(2008C12019),浙江省院合作林业科技项目“早实高产白花树生物柴油能源林定向培育与制取工艺研究”(2010SY 05),浙江省创新团队建设与人才培养项目(2010F20014)
赵勋(1975-),男,安徽蚌埠人,硕士研究生,从事园林植物研究。*通讯作者。
