高温胁迫下牡丹的抗逆生理响应
2011-04-26韩金蓉艳柯费永俊长江大学园艺园林学院湖北荆州434025
骆 俊,韩金蓉,王 艳柯 林,杨 敏,费永俊 长江大学园艺园林学院,湖北荆州434025
牡丹 (Paeonia suf f ruticosa)属芍药科芍药属,多年生木本落叶灌木,是我国特有的观赏植物种质资源,具有重要的观赏、食用、药用及工业价值[1]。近十余年,国内外展开了对牡丹的系统研究,主要集中在资源的调查、品种分类、引种栽培、细胞学、分子生物学、生理等方面。生理方面的研究主要集中在花期调控、切花采后生理和贮藏保鲜[2~6],但关于牡丹抗高温胁迫耐性方面的报道很少。本研究通过研究高温胁迫下牡丹叶片电解质渗透率、游离脯氨酸 (Pro)含量、丙二醛 (MDA)含量和过氧化物酶 (POD)活性等抗性生理指标的变化,研究牡丹抗胁迫的形式、途径和大小及抗性机制,初步探讨牡丹高温胁迫下的伤害与适应等抗性生理变化,以更好地开发利用牡丹资源。
1 材料与方法
1.1 试验材料
试材取自于植物园内种植的一年生牡丹幼苗,种源采自湖北五峰县野生牡丹,已在长江大学植物园驯化栽培8 a,能正常开花结实。于2009年9月3日选取牡丹幼苗移栽入花盆,栽培基质配方为蛭石∶珍珠岩∶泥炭土∶陶粒=3∶1∶4∶1。
1.2 试验处理
选取生长势一致的盆栽牡丹置于人工气候箱内,分别对牡丹进行不同模拟高温处理,设置30、35、40℃3个温度,并以25℃为对照;每种方式处理3盆,处理过程中气候箱内空气湿度为80%,光照强度为4 000 lx,每天光照12 h,连续处理2 d。处理后立即从各苗木基部采取3片叶片进行测定,每个指标重复测定3次。
1.3 指标测定方法
(1)电解质渗透率测定 准确称取不同处理的待测植物叶样0.2 g,分别置于烧杯中,加蒸馏水10 mL,在25℃下浸提30 min后,采用DDS-11A型电导仪分别测定电导率 (C1),再置于沸水浴中10 min,冷却后分别测定煮沸电导率 (C2),然后采用公式电解质渗透率=(C1/C2)×100%[7]分别计算电解质渗透率。
(2)游离脯氨酸含量测定 准确称取不同处理的待测植物叶样各0.2 g,分别置入大试管中,分别加入3%的磺基水杨酸溶液5 mL,在沸水浴中提取10 min(提取过程中要经常摇动),冷却后过滤于干净的试管中,滤液即为脯氨酸的提取液。吸取提取液2 mL于另一干净的带玻塞试管中,加入冰醋酸2 mL及酸性茚三酮试剂2 mL,在沸水浴中加热30 min,溶液即呈红色。冷却后加入甲苯4 mL,摇荡30 s,静置片刻,取上层液至10 mL离心管中,在3 000 r/min下离心5 min。然后用吸管轻轻吸取上层脯氨酸红色甲苯溶液于比色杯中,以甲苯为空白对照,在分光光度计上520 nm波长处比色,测得光密度值X,再根据公式单位鲜重样品的脯氨酸含量=X×2.5/样重[7]计算出脯氨酸含量。
(3)MDA含量的测定 准确称取不同处理的待测植物叶片各0.2 g,加入5%TCA 5 mL和少量石英砂,研磨成匀浆,匀浆在3 000 r/min离心10 min,上清液为样品提取液。吸取离心的上清液2 mL于另一干净试管中,加入0.67%TBA溶液2 mL,混匀物于沸水浴中反应30 min,迅速冷却后再离心1次。取上清液于比色杯中,以清水为对照,在分光光度计上分别测定600、532、450 nm波长下的光密度。用公式 C2=6.45×(D532-D600)-0.56×D450计算MDA的浓度,然后用公式MDA含量(μ mol/g)=C2μ mol/L)×提取液体积 (mL)/植物组织鲜重 (g)计算MDA含量[7]。
(4)POD活性的测定 准确称取不同处理的待测植物叶片各0.2 g,加 20 mmol/L KH2PO42.5 mL于研体中研磨成浆,在4 000 r/min离心15 min,倾出上清液在冷处保存,残渣再加KH2PO42.5 mL提取1次,合并2次的上清液,冷处保存备用。取比色杯2只,1只加反应混合液3 mL、KH2PO41 mL,另1只加3 mL反应混合液、1 mL酶液,在470 nm处读数 (每分钟1次)。以每分钟光密度变化 (以每分钟D470nm变化0.01为1个活力单位)表示酶活性大小[7]。
1.4 数据处理
数据用DPS软件进行统计分析。
2 结果与分析
2.1 高温胁迫下牡丹的形态变化
观察发现,牡丹在25℃条件下能正常生长,在35℃的高温下处理48 h后,牡丹叶片出现轻微失水症状,叶片稍下垂;40℃高温处理后叶片失水、叶片尖端和叶缘出现褐色焦枯色块等症状。
2.2 高温胁迫下牡丹的抗逆生理响应
(1)高温胁迫对牡丹电解质渗透率的影响 如图1所示,牡丹经30、35、40℃高温处理后,质膜透性的变化较大,温度越高质膜透性越大,40℃与35、30、25℃有极显著差异,35℃与30、25℃有显著差异。说明高温对牡丹叶片细胞的伤害是明显的。
(2)高温胁迫对牡丹MDA含量的影响 如图2所示,随着温度的升高,MDA含量总体上呈现先上升后下降的趋势,各处理下的MDA含量均高于对照。可能高温胁迫初期使牡丹叶片内的MDA含量增加,但35℃以上的高温胁迫使牡丹的防御机能出现紊乱,又使MDA含量降低。牡丹各温度处理下叶片中MDA含量均高于对照,说明高温对细胞膜的破坏明显。
(3)高温胁迫对牡丹Pro含量的影响 图3所示高温30、35、40℃处理的 Pro含量分别是 0.013、0.0277、0.031 g鲜重,分别为对照的 39.78%、190.32%、240.86%,且40℃与35、30、25℃均有显著差异,可见,高温胁迫能引起Pro含量的变化,表明随着逆境胁迫程度的增加,牡丹叶片中脯氨酸含量增多,因此,脯氨酸可作为牡丹抗逆性强弱的鉴定指标。

图1 高温胁迫对牡丹电解质渗透率的影响
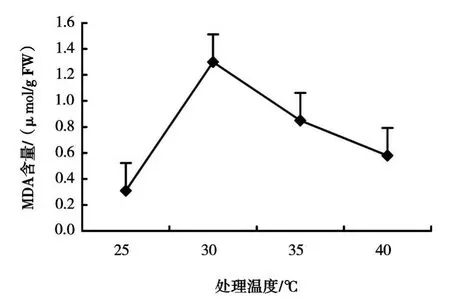
图2 高温胁迫对牡丹MDA含量的影响
(4)高温胁迫对牡丹过氧化物酶 (POD)活性的影响 图4显示出了经寒热胁迫后POD活性的变化情况,与25℃对照相比,40℃与35、30、25℃有极显著性差异,35℃与30℃有极显著性差异,30℃与25℃有显著性差异,随着胁迫温度的升高,牡丹叶片中的POD活性变化总体呈先降后升的趋势。

图3 高温胁迫对牡丹脯氨酸含量的影响

图4 高温胁迫对过氧化物酶(POD)活性的影响
2.3 Pro含量及其与电解质渗透率的相关性
高温胁迫伤害细胞膜,改变膜透性,使胞质外渗量增加,电解质渗透率高,而脯氨酸是一种植物渗透调节剂,其含量的增加有助于细胞持水和生物大分子结构的稳定[8]。将测定的游离脯氨酸 (Pro)含量与电解质渗透率作相关性分析,发现二者之间存在显著性正相关,相关系数为0.985(表1)。

表1 牡丹生理指标间相关系数
2.4 POD活性与MDA的关系
POD是植物对膜脂过氧化作用酶促防御体系的重要保护酶之一,通常POD活性的增加趋势与MDA含量增加的趋势呈负相关[9,10]。由表1可见,POD活性与MDA含量呈负相关,相关系数为-0.3357,达到显著水平。
3 讨论与结论
3.1 高温胁迫与电解质渗透率、游离脯氨酸 (Pro)的关系
生物膜在植物逆境胁迫研究中占有重要地位,在高温胁迫下,植物细胞膜系统的半透性逐步丧失,依赖于膜系统的代谢系统和信号传递系统发生紊乱。当牡丹受到高温胁迫时,电解质渗透率都有所增加,并随着高温胁迫强度的增强而继续增加。
逆境下植物叶片游离脯氨酸累积,一是因为叶片组织中多种酶活性降低,脯氨酸氧化受阻,造成游离脯氨酸积累;二是谷氨酸合成脯氨酸的速度增加。脯氨酸是一种植物渗透调节剂,可提高植物细胞原生质渗透压,防水分散失以及提高原生质胶体的稳定性,从而提高植物体抗性[11]。大量的研究结果表明,脯氨酸的积累与细胞的脱水有关,由于胁迫引起脱水,因此发生脯氨酸的积累。它的积累除可起到渗透调节外,更重要的是对膜脂和蛋白起到保护作用,防止活性氧对膜脂和蛋白的过氧化作用[12]。由此可以认为高温胁迫下牡丹叶片内脯氨酸的升高也有助于提高它的抗性。
在逆境下植物体内常有游离脯氨酸的积累,其积累量与逆境水平和植物对这种逆境的抗性有关,因而测定植物体内游离脯氨酸含量在一定程度上可以了解植株遭受逆境的情况及植物对逆境的抵抗能力。从本研究结果来看,随着高温程度的加剧,牡丹叶片游离脯氨酸的累积随高温时间呈指数上升趋势,而且品种与品种之间表现出明显的差异。连续高温使植株受损伤的程度大,体内游离脯氨酸积累量大。因此,游离脯氨酸可作为牡丹高温胁迫的可靠指标。
3.2 高温胁迫与过氧化物酶 (POD)的关系
POD是膜保护系统的组成成分之一,能在逆境中清除植物体内的活性氧,维持体内的活性氧代谢平衡,保护膜结构,减轻有毒物质对活细胞的毒害,延迟和阻止细胞结构的破坏,使组织保持活力,从而使植物在一定程度上忍耐抵抗高温逆境胁迫。图4所示,温度由25℃升至35℃时,酶活性缓慢下降,温度继续升高时,酶活性显著上升。表明POD对环境条件的变化敏感,高温胁迫时POD活性降低,可能是高温胁迫打乱了植物正常的生理机能,暂时的不适应导致了POD的合成受阻,一段时间后,它们又开始积极的防御反应。
3.3 高温胁迫与丙二醛 (MDA)含量的关系
MDA是高活性的脂过氧化物,能交联脂类、核酸、糖类及蛋白质,在细胞中的积累具有很强的细胞毒性,常导致质膜伤害。叶片中MDA含量的多少可以代表膜损伤程度的大小。由于高温胁迫,植物器官衰老或受到伤害,细胞往往发生膜质过氧化作用,丙二醛是膜质过氧化的最终分解产物,细胞内活性氧代谢的平衡被破坏,从而导致活性氧的产生,活性氧的毒害之一是引发或加剧膜脂过氧化作用,造成细胞系统的损伤,因而作为膜脂过氧化产物,MDA含量的变化是质膜损伤程度的重要标志之一。因此MDA含量可以反映植物遭受高温胁迫伤害的程度。
3.4 高温胁迫下POD活性与MDA含量的关系
POD活性与MDA含量成显著负相关 (表1),这种现象一方面是牡丹在高温胁迫下活性氧累积加剧了膜脂过氧化作用,导致体内MDA的积累,而MDA积累量还未达到能反过来抑制POD等保护酶的活性的程度,即活性氧的积累水平还处在膜脂过氧化作用酶促和非酶促防御体系所能调控的阈值范围内[13,14]。另一方面与牡丹叶片POD在高温胁迫中反应的复杂性和作用的非专一性有关。
4 结论
正常情况下,植物体内各项代谢的生理生化过程都是比较稳定而协调的,当植物受到逆境胁迫时,植物体内的各种代谢活动都会由于某种因素的影响而失调,使植物对逆境作出反应。本研究结果表明,当牡丹开始受到高温胁迫时,牡丹叶片的电导率增大,游离脯氨酸大量累积,过氧化物酶活性升高,牡丹能正常生长,这说明牡丹具有一定的抗热性,即能通过一系列保护性的生理生化反应来适应高温胁迫,以减轻伤害;但随着胁迫温度的增强,牡丹的外部形态和生理变化较大。
[1]王莲英,袁 涛.国牡丹与芍药[M].北京:金盾出版社,1999.
[2]李莹莹.我国盆栽牡丹研究进展与发展动向 [J].潍坊教育学院学报,2010,23(1):71-73.
[3]赵一鹏,蔡祖国,李本勇.珍稀濒危植物矮牡丹研究进展 [J].河南农业科学,2009,(7),14-17.
[4]周秀梅,李保印.中国牡丹讶花保鲜研究进展[J].北方园艺,2007,(12):60-62.
[5]李保印,周秀梅,蒋细旺,等.牡丹种及品种亲缘关系的分子生物学研究进展 [J].河南农业大学学报,2008,42(1):121-126.
[6]张修景.菏泽牡丹的研究进展 [J].菏泽学院学报,2007,29(5):101-103.
[7]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.182-185.
[8]汤章成.逆境条件下植物体脯氨酸的累积及可能意义[J].植物生理学通讯,1984,(1):15-27.
[9]陈少裕.膜脂过氧化对植物细胞的伤害[J].植物生理学通讯,1999,27(2):84-90.
[10]杜 峰.低温胁迫下草坪草Perennial ryegrass(cv.Taya)和Meadow bluegrass(cv.Compact)各抗性生理指标的变化 [J].四川草原,1998,(3):41-48.
[11]郑小林,胡木林,罗晓莹,等.假俭草低温胁迫的伤害与适应[J].草业科学,2002,19(7):55-57.
[12]Qian Y L,Ball S,Tan Z,et al.Low temperature tolerance of six cultivars of buffalograss[J].Crop Sci,2001,41:1174-1178.
[13]任雪萍,吴诗光,牛明功.湿涝害对灌浆期冬小麦叶片脯氨酸含量的影响 [J].周口师专学报,1998,15(5):165-169.
[14]王翼川,李志军,徐雅丽,等.低温胁迫对海岛棉幼苗生理生化特性的影响[J].中国棉花,2001,28(5):13-14.
