物候分化对天然草地群落生产力的影响
2011-04-25杨志华肖洪浪刘钟龄
赵 亮,王 芳,杨志华,肖洪浪,刘钟龄
(1.中国科学院内陆河流域生态水文重点实验室,甘肃 兰州 730000;2.锡林浩特气象局,内蒙古 锡林浩特 026000;3.内蒙古大学生命科学学院,内蒙古 呼和浩特 010021)
草地是干旱半干旱区生态系统重要的组成部分,对我国黄土高原畜牧业生产具有重要的作用。近年来,过度放牧等人类活动使得天然草地遭到了严重的破坏,影响了天然草地的生态系统服务功能,例如生物多样性丧失、生产力衰退、水土流失加剧、土地沙化等[1]。因此,退化草地恢复重建得到了许多研究者[2-4]的关注,从群落生境、空间结构、植物种类组成、资源状况等多个方面对草地受损恢复进行了研究。草地恢复是草地退化的逆过程,在草地恢复过程中,伴随着生物多样性逐渐提高,从而生态系统的生产力也随之恢复[5]。
生物多样性与生产力的正效应被解释为生态位分化导致互补效应和取样效应的共同作用[6-7]。尽管有许多研究关注生态位分化对初级生产力的影响,然而群落中物种物候期分化,即时间生态位分化,却极少有人关注[8]。草地生态系统物候期分化是广为人知的,物候分化对草地群落多样性维持具有重要的作用[9-10]。然而,物候期分化对初级生产力的影响尚没有明确的结论[11-12]。
为了测定草地群落退化恢复过程中物候期分化对草地生态系统初级生产力的影响,在内蒙古天然草地群落中进行为期两年的试验。与大多数控制试验不同,本研究在物种丰富度较高的天然草地群落中进行,多样性梯度是由于退化草地围封时间不同自然形成的,代表了退化恢复过程中的自然状况。
1 材料与方法
1.1研究区概况 研究地点位于内蒙古自治区锡林河盆地(116°42′ E, 43°83′ N)。天然植被群落的建群种为大针茅(Stipagrandis)和羊草(Leymuschinensis),属于横跨欧亚大陆分布最广泛的天然草地群落。试验地点共包括3块样地,样地都用围栏围封以防止大型食草动物的进入。按照围封时间可分为:样地1,围封于2000年;样地2,围封于1997年;样地3,围封于1983年。样地围封时该地点都因为过度放牧而处于严重退化状态。这3块样地由于围封年代的不同而处于恢复演替的不同阶段,生物多样性沿着恢复时间产生了自然变化梯度。
1.2试验设计 样地从围封后每年都进行8次测定,从5月中旬到9月中旬每15 d测定一次。为了代表不同的演替阶段,本研究选取了2003年和2004年的测定数据,此时样地1处于恢复早期,样地2处于恢复中期,样地3处于恢复后期,组成了一个完整的恢复时间序列。样方内的所有植物按照物种记录株丛数、高度、物候期,地上部分沿地表剪下,按照物种分别装袋,在80 ℃下烘干12 h至质量不变。
初级生产力用地上生物量来代表。本试验中共采用了两种表示方法。第一种是把每年生长期末群落生产力最高时的地上生物量作为样地的初级生产力,这种方法在以草地群落为研究对象的试验中被广泛采用[13];另一种方法是把每个样地的每个物种每年的最大生物量相加作为年净初级生产力(total annual net primary production,TANPP),这种取样方法考虑到了物候期分化产生的互补效应[11-12]。为了不至于产生混淆,本研究中用Bmax来表示群落每年最大地上生物量。
1.3参数选择与统计分析 多样性指数同时采用物种丰富度(S)和物种均匀度(E)两个指标。物种均匀度的计算公式为[14]:
式中,Pi为第i个物种的生物量在TANPP中所占的比例,S为物种丰富度。物候分化程度(Dmax)用超产(overyeilding)程度来表示[15]:
当群落中不存在物候分化时,即所有物种对资源吸收利用都是同步的,那么TANPP=Bmax,Dmax值为0;如果物种之间对资源吸收利用存在物候上的分化,那么群落最大生物量(Bmax)在任何时候都小于TANPP,即TANPP>Bmax,Dmax>0,各物种之间物候期分化的互补效应越大,则Dmax的值越大。由于不能判断研究数据是否符合正态分布,故采用更为稳妥的非参数Spearman相关检验。
2 结果与分析
2.1天然草地群落中的物候分化模式 为了方便图示,采用单个物种在当期群落生产力中所占的比例来代替生物量的绝对值。从图1中可以看出,在内蒙古天然草地群落主要物种之间存在明显的物候期分化的现象。物候期的分化意味着不同物种对资源需要的高峰期之间存在时间上的差异,也就是存在时间生态位的分化。理论上已经证明生态位分化可以提高生态系统的资源利用效率,从而可以提高生态系统初级生产力[15-16]。这也就意味着天然草地群落可以通过物候期分化从自然界获取更多的资源,从而增加系统的初级生产力。
2.2物候期分化与生产力 在所有的样地中,Dmax的值始终大于0,这意味着恢复早期尽管生物多样性水平不高,但由物候期分化造成的超产是天然草地群落中的普遍现象。2003年群落的TANPP与Dmax之间存在显著正相关关系(P<0.001);2004年TANPP与Dmax之间存在显著正相关关系(P=0.001)(图2)。这表明,随着退化恢复时间的进行,物候期分化程度的增加提高了群落对资源的利用效率,从而增加了生态系统的初级生产力。

图1 天然草地群落物候分化示意图(以2004年样地3为例)
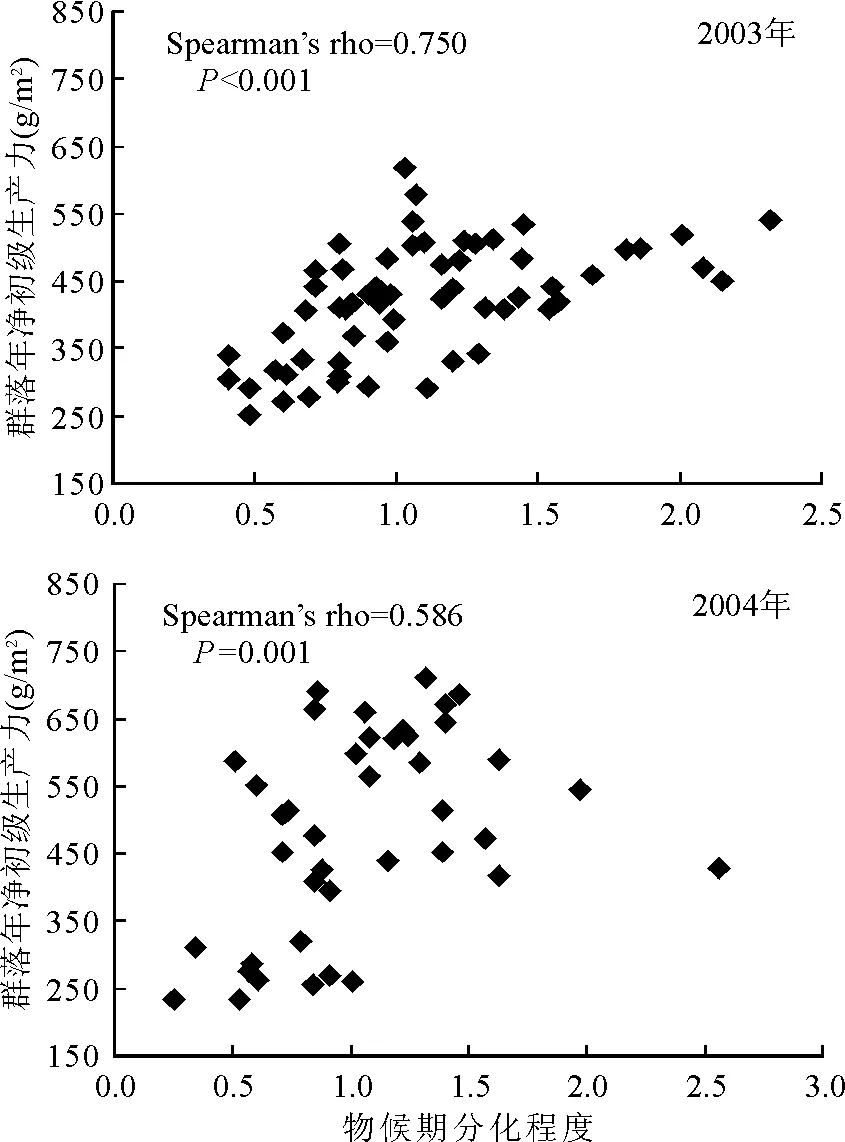
图2 物候期分化与群落年净初级生产力之间的关系
2.3物候期分化与生物多样性 进一步分析物候期分化与生物多样性之间的关系,发现在两年的研究期内,群落的物种丰富度和物种均匀度与物候期分化之间均呈显著的正相关关系(图3)。随着退化草地的恢复,群落中出现的物种数目开始增加,占据不同生态位的物种逐渐恢复到群落中,或者某些物种在群落中的数目增加,改变了群落的物种均匀度。而物种丰富度和均匀度的增加都能提高资源利用效率从而提高群落生产力。
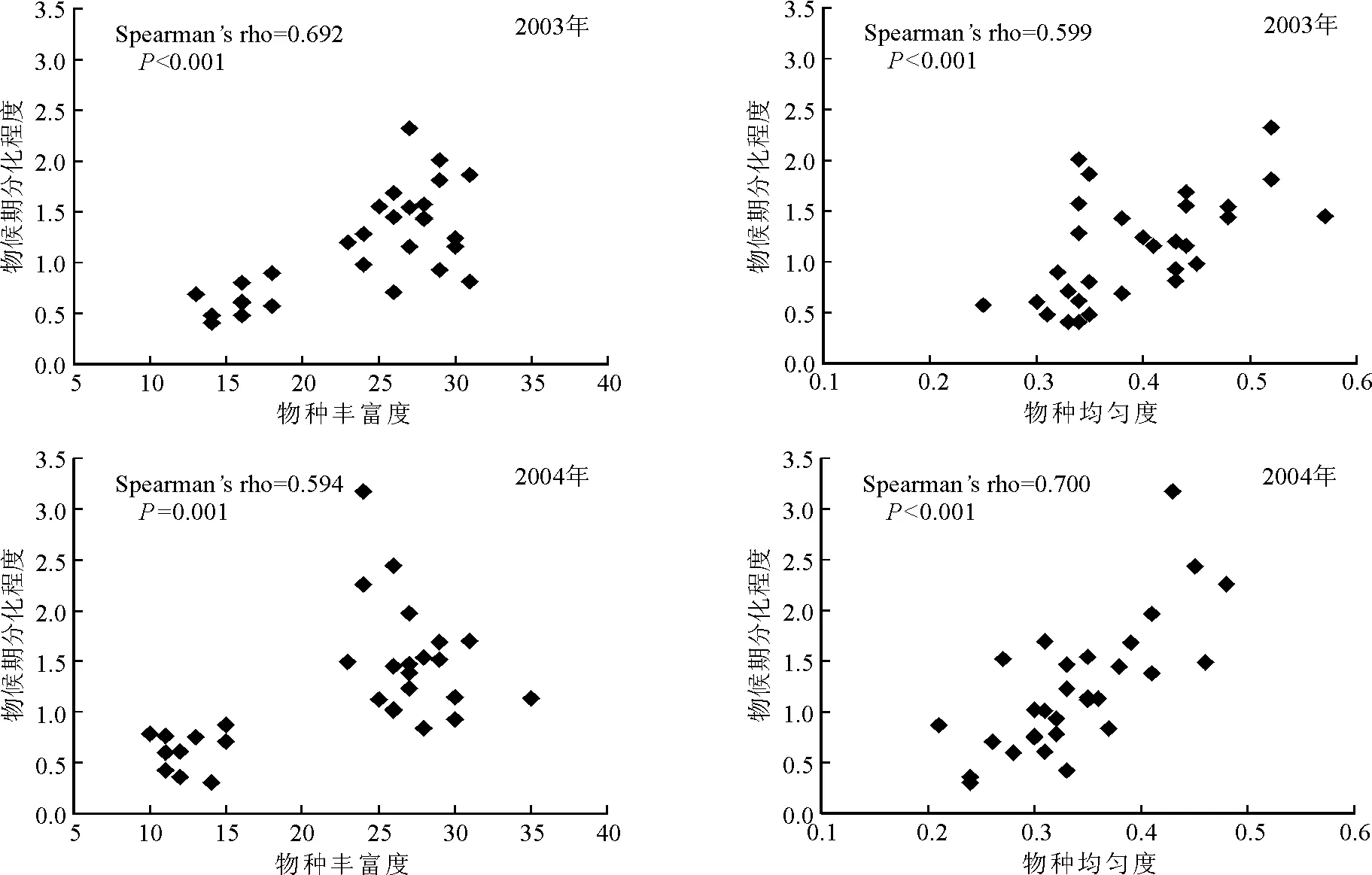
图3 物候期分化程度与物种丰富度、物种均匀度的关系
3 讨论
锡林郭勒草原在20世纪八、九十年代退化十分严重,到围栏封牧时已因过度放牧退化成以冷蒿(Artemisiafrigida)为优势种的群落类型,主要以一年生的旱生、中旱生植物为主[3]。放牧扰动没有改变该地区群落的物种库,只是引起了一些种群数量的改变,土壤养分等资源与原生群落相比并没有显著的差异,放牧压力使得一些生态位不能被占据[2]。围栏后放牧压力消失,随着恢复时间的延长,这些空出来的生态位被重新占据。本研究表明,群落在时间生态位维度方面也存在生态位被重新占据的现象,即物候期分化程度更加显著,物候期分化通过互补效应对生态系统初级生产力具有显著的促进作用。不考虑时间生态位的分化可能会低估生物多样性对生态系统功能的影响。
大多数被子植物的物候期与生长量密切相关,生长量则是植物从环境中获取物质和能量的体现。植物体所需要的养分十分相近,在生物多样性高的群落中,物候期分化程度可能对群落中多个物种共存具有重要的意义[10,17]。植物物候期的变化与光照、温度等环境因子密切相关,植物物候期对环境因子的响应种间差异很大。近年来的一些研究[18-20]表明,全球气候变化正在导致植物群落物候期的改变,因此物候期分化对植物群落生态系统服务功能的影响将成为未来研究的一个潜在的重要方向。
[1]赵艳云,胡相明.黄土高原退化草地恢复演替进展研究[J].水土保持研究,2008,15(6):270-272.
[2]王炜,刘钟龄,郝敦元,等.内蒙古草原退化恢复演替的研究Ⅰ.退化草地的基本特征与恢复演替动力[J].植物生态学报,1996,20(5):449-459.
[3]刘钟龄,王炜,郝敦元,等.内蒙古草原退化与恢复演替机理的探讨[J].干旱区资源与环境,2002,16(1):84-87.
[4]魏永胜,梁宗锁,山仑.草地退化的水分因素[J].草业科学,2004,21(10):13-18.
[5]Hooper D U,Chapin III F S,Ewel J J,etal.Effects of biodiversity on ecosystem functioning: a consensus of current knowledge[J].Ecological Monographs,2005,75:3-35.
[6]Huston M A.Hidden treatments in ecological experiments: re-evaluating the ecosystem function of biodiversity[J].Oecologia,1997,110:449-460.
[7]Loreau M S,Hector A.Partitioning selection and complementarity in biodiversity experiments[J].Nature,2001,412:72-76.
[8]Zhao L,Yang G X,Liu Z L,etal.Phenological complementarity doesn’t enhance ecosystem production in undisturbed steppe community:a three years case study[J].Journal of Integrative Plant Biology,2007,49(5):582-587.
[9]Rathcke B,Lacey E P.Phenological patterns of terrestrial plants[J].Annual Review of Ecology and Systematics,1985,16:179-193.
[10]杜国祯,赵松岭.草本群落中物种多样性维持机制——花期不同步性[J].草业学报,1997,6(3):1-7.
[11]Hooper D U.The role of complementarity and competition in ecosystem responses to variation in plant diversity[J].Ecology,1998,79:704-719.
[12]Hooper D U,Dukes J S.Overyielding among plant functional groups in a long-term experiment[J].Ecology Letters,2004,7:95-105.
[13]van Ruijven J,Berendse F.Diversity-productivity relationships: initial effects,long-term patterns,and underlying mechanisms[J].Proceedings of the National Academy of Sciences of the United States of America,2005,102:695-700.
[14]Smith M D,Wilson J B.A consumer’s guide to evenness indices[J].Oikos,1996,76:70-82.
[15]Loreau M.Biodiversity and ecosystem functioning:a mechanistic model[J].Proceedings of the National Academy of Sciences of the United States of America,1998,95:5632-5635.
[16]Loreau M.Biodiversity and ecosystem function:recent theoretical advances[J].Oikos,2000,91:3-17.
[17]李向前,贾鹏,章志龙,等.青藏高原东缘高寒草甸植物群落的开花物候[J].生态学杂志,2009,28(11):2202-2207.
[18]Fenner M.The phenology of growth and reproduction in plants[J].Perspectives in Plant Ecology, Evolution and Systematics,1998,1:78-91.
[19]Crawley M J.Timing of disturbance and coexistence in a species-rich ruderal plant community[J].Ecology, 2004,85:3277-3288.
[20]张峰,周广胜,王玉辉.内蒙古克氏针茅草原植物物候及其与气候因子关系[J].植物生态学报,2008,24(5):1312-1322.
