饲粮能量水平对乌金猪脂肪组织脂类合成代谢相关基因表达的影响
2011-03-28潘洪彬赵素梅葛长荣高士争
潘洪彬 赵素梅 黄 英 王 静 张 曦 葛长荣 高士争
乌金猪是云南省优良的地方猪种之一,具有耐粗饲、抗逆性强、肉质优良、肌内脂肪丰富等优良性状[1]。研究不同能量水平的饲粮对乌金猪脂肪组织中脂类合成代谢和脂肪酸转运基因表达的影响,对阐明饲粮能量水平对猪脂肪组织脂类代谢影响的分子机制具有重要科学意义。
动物体脂的沉积是一个复杂生理生化过程,受动物品种、年龄及营养状态的影响较大。在脂肪合成过程中,乙酰辅酶 A羧化酶(acetyl CoA carboxylase,ACC)和脂肪酸合成酶(fatty acid synthase,FAS)催化乙酰辅酶 A合成脂肪酸[2-4],是脂肪合成的限速酶[5-6]。固醇调节元件结合蛋白(sterol regulator element binding protein,SREBP)是一类膜结合转录因子[7],包括 SREBP-la、SREBP-lc和 SREBP-2 3种亚型,营养状态改变可调节SREBP-1c表达[8]。脂肪细胞型脂肪酸结合蛋白(adipocyte fatty acid-binding protein,A-FABP)与肌内脂肪含量、背膘厚及生长发育有关[9-10]。前期试验中,本试验室系统研究了饲粮不同能量水平对乌金猪生长、胴体组成和肉品质的影响,通过模糊综合评定得出了乌金猪在不同生长阶段获得最佳肉品质的所需的饲粮能量水平[11-12],并研究了饲粮蛋白质对猪的脂肪组织中脂类代谢相关基因表达的影响[13]。本试验在此基础上,进一步深入研究饲粮不同能量水平对乌金猪脂肪组织脂类合成代谢相关基因表达的影响,阐明饲粮能量水平对猪脂肪组织脂类合成代谢影响的分子机制。
1 材料与方法
1.1 试验动物及试验设计
选取健康、胎次相近、体重约 15 kg的乌金猪54头,公母各占 1/2,采用单因子随机分组分为 3组,每组 3个重复,每个重复 6头猪,分别饲喂 3种不同消化能水平的饲粮。
根据《动物营养参数与饲养标准:中国肉脂型生长肥育猪饲养标准》[14]设计乌金猪各生长阶段饲粮,每阶段饲粮粗蛋白质等营养水平基本一致,设低(LDE)、中 (MDE)、高(HDE)3个消化能水平,中能组饲粮采用饲养标准的消化能,低、高能组饲粮则分别上下调整 10%,消化能分别为低能组 11.74 MJ/kg、中能组 12.89 MJ/kg、高能组14.22 MJ/kg[11-12]。试验饲粮组成及营养水平见表1。
1.2 饲养管理
试验分 15~30、30~60和 60~100 kg 3个阶段。预试期为 5 d,试验期日饲喂 3次,自由饮水,注意观察猪的精神状态和行为等,各阶段换料采用逐渐过渡方法,过渡期为 7 d。分别于 30、60和100 kg体重时分批屠宰,每批屠宰 6头,测定胴体脂肪率,并取皮下脂肪组织于 -80℃保存待测。
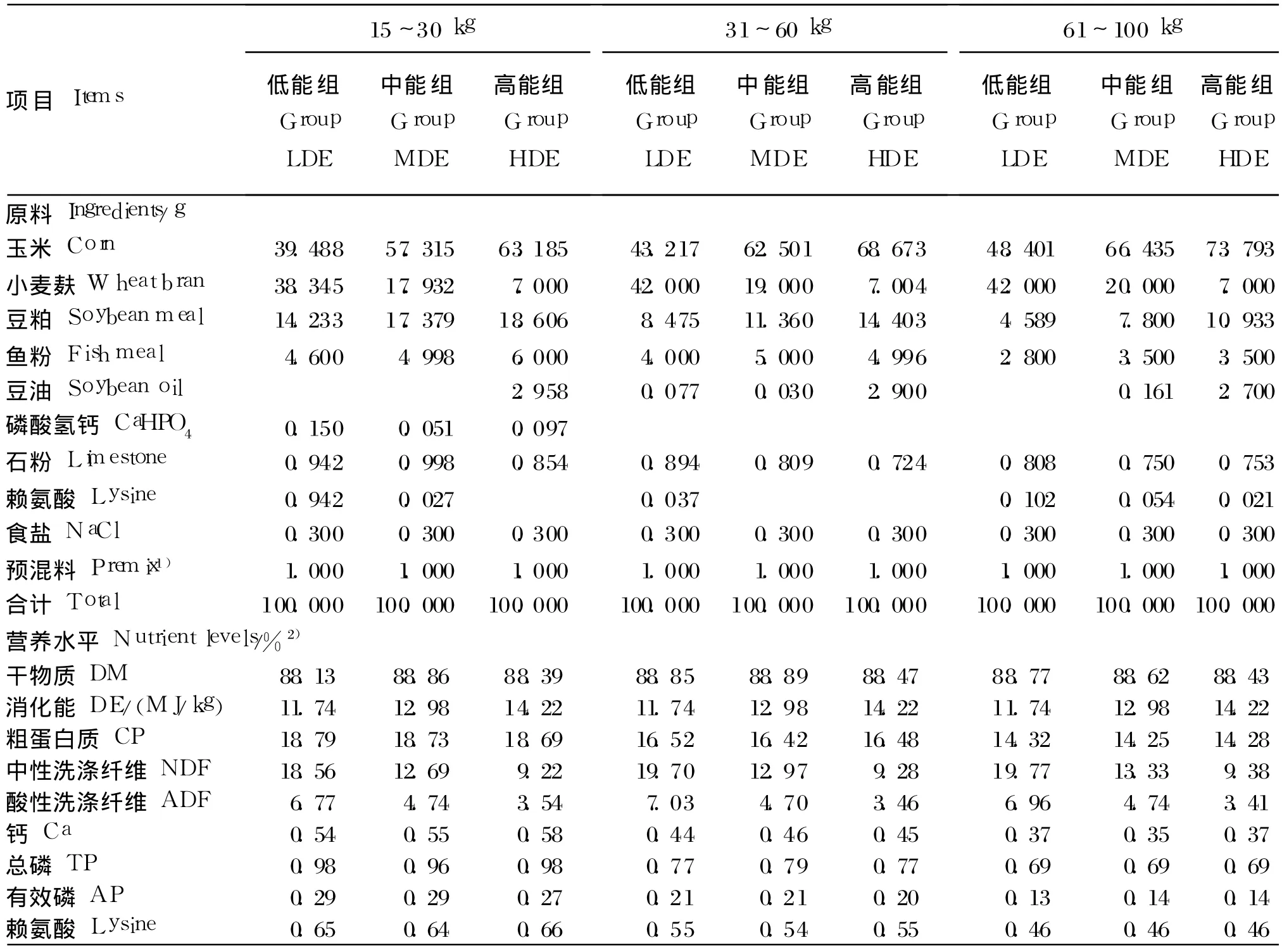
表1 试验饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of experimental diets(air-dry basis)
1.3 实时定量 PCR(real-time PCR)
1.3.1 总 RNA提取
用 Trizol试剂提取总 RNA,用紫外分光光度计(260 nm)和变性琼脂糖凝胶电泳测定总 RNA浓度与纯度。
1.3.2 反转录
取 2μg总 RNA进行反转录,反应采用 20μL体系:12μmol/L随机引物、0.5 mmol/L dNTP、20 U/μL RNA酶 抑 制 剂 (RNase inhibitor)、10 U/μL反转录酶(M-MLV RTase)、4μL 5×RT Buffer[含 250 mmol/L Tris-HCl(pH 8.3)、50 mmol/L MgCl2、 250 mmol/L KCl、50 mmol/L二硫苏糖醇(DTT)、2.5 mmol/L亚精胺(Spermidine)]。先加 RNA模板、dNTP和随机引物,70℃变性 5 min,立即冷却,再加其余试剂 37℃反应60 min,95℃灭活 5 min。反转录产物于 -20℃保存备用。
1.3.3 实时定量 PCR
目的基因(ACC、FAS、SREBP-1c、A-FABP)和内标基因 β-肌动蛋白(β-actin)引物使用 Primer v5软件根据 GenBank上猪的基因序列设计,引物序列、退火温度、产物大小及登录号见表 2。

表2 实时定量PCR特异性引物Table 2 Specific primers for real-time PCR
实时定量 PCR采用 20μL反应体系:1.5μL反转录产物、10μL iQTMSYBRGreen Supermix(BIO-RAD,美 国 )、 0.25 mmol/μL dNTP、0.5 mmol/L目的基因引物。每个样品测定 2次,2个平行样间的相对偏差不大于 10%。各基因 PCR的优化条件见表 3。

表3 实时定量 PCR条件Table 3 Conditions for real-time PCR
1.3.4 定量方法
实时定量 PCR结果以表达量的倍数表示,基因表达量根据下列公式计算[15]。
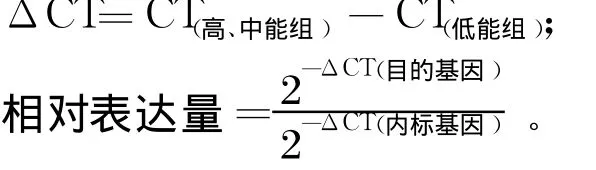
式中,CT为阈值循环。
1.4 数据分析
采用 SAS 9.0软件对试验数据进行统计分析,对平均值进行方差分析,做显著性检验(P<0.05、P<0.01)。
2 结 果
2.1 饲粮能量水平对不同生长阶段乌金猪脂肪率的影响
由表 4可知,不同能量水平的饲粮对 30 kg乌金猪脂肪率影响不显著(P>0.05),而对 60和100 kg的乌金猪的脂肪率有极显著影响(P<0.01),高能组乌金猪的脂肪率比低能组的分别高8.39%和 19.25%,且乌金猪脂肪率随饲粮能量水平的升高呈现升高趋势。
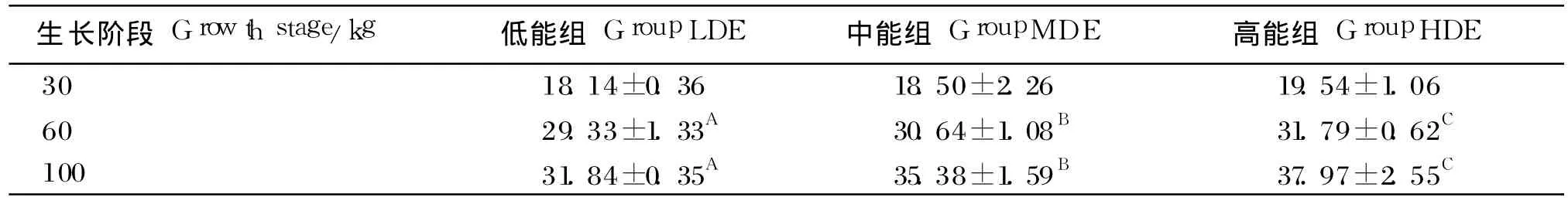
表4 不同生长阶段饲喂不同能量水平饲粮的乌金猪的脂肪率Table 4 Fat percentage of Wujin pigs feed diets with different energy levels at different growth stage %
2.2 饲粮能量水平对不同生长阶段乌金猪 ACC表达水平的影响
由表 5可知,体重为 30 kg时,中、高能组脂肪组织 ACC表达水平分别比低能组降低了65.00%、78.00%(P<0.05),体重为 60 kg时,分别提高了 4.73%(P>0.05)和 24.54%(P<0.05),体重为 100 kg时,分别降低了 3.43%(P>0.05)和32.31%(P<0.05)。此外,体重为 60 kg时,脂肪组织 ACC基因的表达水平高于体重为 30和100 kg时。
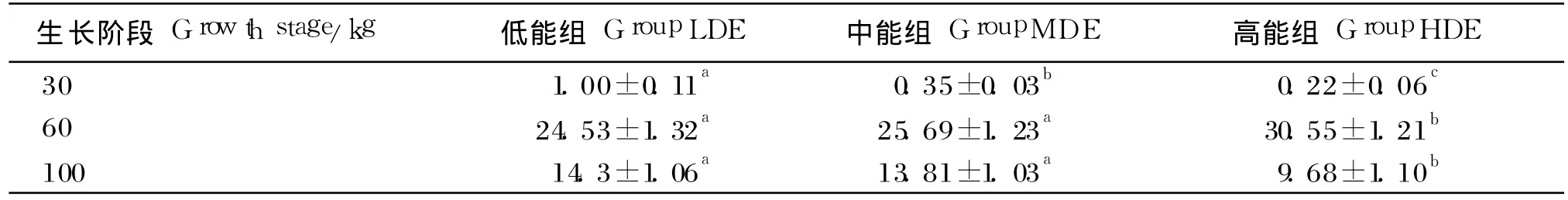
表5 不同生长阶段饲喂不同能量水平饲粮的乌金猪脂肪组织ACC表达水平Table 5 The expression level of ACC in adipose tissue of Wujin pigs feed diets with different energy levels at different growth stage
2.3 饲粮能量水平对不同生长阶段乌金猪 FAS表达水平的影响
由表 6可知,60 kg体重时,脂肪组织 FAS表达水平随饲粮能量水平升高而升高,中、高能组分别比低能组的升高了 79.56%和 227.73%(P<0.05);在 30和 100 kg体重时则相反,FAS表达水平随饲粮水平的升高而降低,中能组比低能组的分别降低 11.00%和 3.92%(P>0.05);高能组比低能组分别降低了 65.00%和 38.24%(P<0.05)。此外,60 kg体重时,脂肪组织 FAS基因的表达水平比 30和 100 kg体重时高。
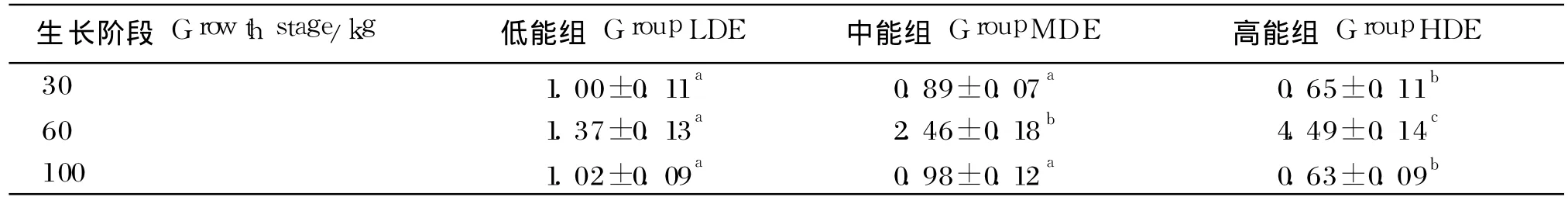
表6 不同生长阶段饲喂不同能量水平饲粮的乌金猪脂肪组织 FAS表达水平Table 6 The expression level of FAS in adipose tissue of Wujin pigs feed diets with different energy levels at different growth stage
2.4 饲粮能量水平对不同生长阶段乌金猪SREBP-1c表达水平的影响
由表 7可知,高能组与低、中能组脂肪组织SREBP-1c基因表达水平差异显著(P<0.05);体重为 60 kg时,脂肪组织 SREBP-1c表达水平随饲粮能量水平的升高而升高,高能组与中能组相比显著上调 62.07%(P<0.05);体重为 30和100 kg时,SREBP-1c表达水平随饲粮能量水平的升高而降低,中能组比低能组分别降低 5.00%和3.03%,高能组比低能组分别降低 45.00%和 24.24%。并且,体重为 60 kg时,脂肪组织 SREBP-1c表达水平比 30和 100 kg体重时高。
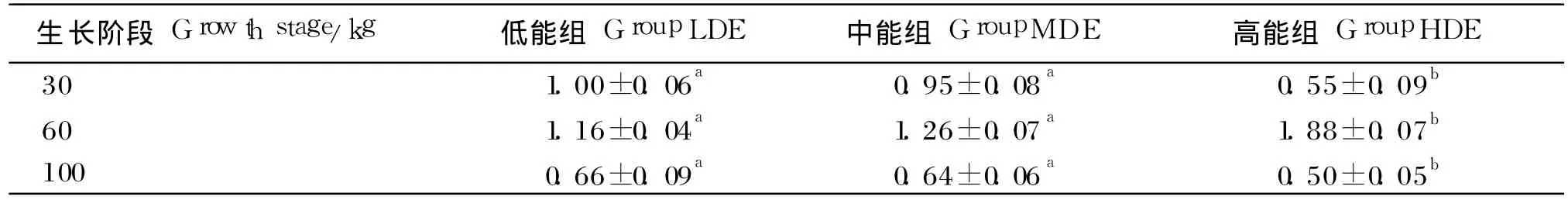
表7 不同生长阶段饲喂不同能量水平饲粮的乌金猪脂肪组织 SREBP-1c表达水平Table 7 The expression level of SREBP-1c in adipose tissue of Wujin pigs feed diets with different energy levels at different growth stage
2.5 饲粮能量水平对不同生长阶段乌金猪A-FABP表达水平的影响
由表 8可知,随着饲粮能量水平的升高,30和100 kg体重时脂肪组织 A-FABP表达水平逐渐升高,中能组分别比低能组显著升高 119.00%和65.00%(P<0.05),高能组分别比低能组显著升高 339.00%和 120.00%(P<0.05);体重为60 kg时,A-FABP表达水平随饲粮能量水平的升高而降低,中、高能组分别比低能组显著降低了 20.45%和 67.05%(P<0.05)。此外,随着体重的增加,除高能组外各组 A-FABP表达有逐渐下降的趋势。

表8 不同生长阶段饲喂不同能量水平饲粮的乌金猪脂肪组织 A-FABP表达水平Table 8 The expression level of A-FABPin adipose tissue of Wujin pigs feed diets with different energy levels at different growth stage
3 讨 论
动物每天从食物中摄取能量,并主要在肝脏和脂肪组织中把多余的能量转变成脂肪储存起来。能量供给水平与畜禽的生产性能和胴体品质有密切关系,一般来说,畜禽摄取能量越多,日增重越快,饲料利用率越高,胴体脂肪含量也越多。研究表明,提高饲粮能量水平可促进生长,同时又增加体脂含量[15-17],刘作华等[18]研究也表明饲粮能量水平影响猪生产性能及体脂沉积。本试验结果表明,不同生长阶段的乌金猪脂肪率随饲粮能量水平的升高而升高,与上述研究结果相一致。
中国地方猪种与国外其他商品猪种相比,表现生长速度慢、脂肪沉积率高[19-22]。脂肪沉积主要与脂肪组织的脂肪生成能力有关,脂肪组织是脂肪酸合成的主要部位[23],脂肪和肌肉组织中甘油三酯的数量是脂肪合成和降解的综合体现。熊文中等[24]研究发现,猪脂肪组织中脂肪酸合成酶与胴体脂肪量、胴体的脂肪率呈极显著正相关。另外,动物脂肪的水解主要靠激素敏感脂酶催化水解,这些酶是脂肪细胞中甘油三酯降解的限速酶,在调控能量平衡中起重要作用[25]。
ACC是脂肪酸从头合成过程中的关键酶[2],它调节脂肪酸合成以及脂肪酸的 β氧化,营养因素对 ACC活性及其 mRNA水平的调控具有组织特异性,ACC在脂肪型猪体内的活性是瘦肉型猪的 3倍,它与基础脂肪的合成率呈正相关[20];FAS是一个多酶复合体系,它利用丙二酸单酰辅酶 A合成长链脂肪酸棕榈酸[5],FAS基因的表达增加能显著促进甘油三酯的沉积从而导致肥胖[26]。刘作华等[18]研究表明能量水平可通过影响 FAS的表达来调节猪的脂肪沉积。此外,ACC和 FAS对脂肪的合成起到限速性和决定性的作用[5-6]。SREBP-1c是一类与膜结合的转录因子,它们能够激活编码胆固醇和脂肪酸生物合成的酶基因,这些酶包括 ACC、FAS、硬脂酰辅酶 A去饱和酶、甘油 -3-磷酸乙酰转移酶等[27-28]。本试验结果表明,高能量饲粮显著促进 60 kg体重乌金猪脂肪组织中 ACC、FAS和 SREBP-1c基因的相对表达,脂肪合成相关的基因表达受到上调,说明 60 kg乌金猪脂肪组织中的游离脂肪酸从头合成有所增加,相反高能量饲粮却显著抑制 30和 100 kg体重乌金猪脂肪组织中 ACC、FAS和 SREBP-1c基因的表达,间接说明高能量饲粮降低了乌金猪生长早期和后期的脂肪组织中脂肪酸的从头合成。
脂肪组织中 A-FABP负责将脂肪细胞的脂肪酸运送至脂肪酸氧化或三酰甘油合成的位置[29],防止细胞内堆积过多的游离脂肪酸,并维持脂肪细胞脂解和脂肪合成之间代谢的动态平衡[30]。本试验结果表明,高能量饲粮促进了 100 kg体重乌金猪 A-FABP的基因表达,并且 100 kg体重时乌金猪胴体脂肪率明显高于其他体重阶段,说明脂肪酸的转运增强,脂肪组织摄取较多的游离脂肪酸,脂肪合成能力增强;而 30 kg体重乌金猪的A-FABP的基因表达增强,也许是生长早期需要储备一定量的脂肪以利于中后期的生长;高能量饲粮抑制 60 kg体重乌金猪脂肪组织 A-FABP基因表达,说明脂肪组织对外源脂肪酸的利用减少,即分解利用的脂肪酸减少,而体内从头合成的脂肪酸增加;从乌金猪脂肪率随饲粮能量水平升高而升高的趋势来看,乌金猪机体随能量水平的升高可能储存了更多的脂肪酸,即高能量饲粮能增加脂肪组织中的脂肪酸含量。
因此,乌金猪在不同生长阶段,饲粮能量水平对脂肪组织的脂肪代谢可能存在不同的调控机制,在生长早期和后期(30和 100 kg),饲粮高能量抑制了脂肪组织中脂肪酸的从头合成,但高能量水平却能增加脂肪组织的脂肪沉积[15-17],说明高能量营养有可能通过其他脂类合成途径合成脂肪,增加脂肪沉积,本试验结果也证实了此时脂肪酸的转运增加,说明脂肪组织摄取了较多的外源脂肪酸来合成脂肪,可能通过二脂酰甘油酰基转移酶(DGAT)来对外源脂肪酸加以利用,合成甘油三酯[31]。而生长中期(60 kg)的情况有所不同,此时是体组织合成的旺盛时期,机体蛋白质及脂类代谢都较强,高能量营养同时增强了脂肪酸的从头合成代谢以满足机体快速生长及沉积脂肪的要求。
4 结 论
①体重为 60和 100 kg时,乌金猪脂肪率随饲粮能量水平的升高而极显著升高。
②高能量饲粮显著促进乌金猪脂肪组织脂类合成代谢相关基因(ACC、FAS、SREBP-1c)在体重60 kg时的表达,可能增加了 60 kg体重乌金猪脂肪组织中游离脂肪酸的从头合成。
③高能饲粮显著抑制脂肪酸转运相关基因(A-FABP)在 60 kg时的表达,降低了脂肪酸转运,可能减少了脂肪组织对外源脂肪酸的摄取。
[1] 王忠庆,荣耀先.大河乌猪与大河猪(乌金猪)肉质形状研究[J].云南农业科技,2004,增刊:41-44.
[2] LIU C Y,GRANT A L,KIM K H,et al.Porcine somatotropin decreases acetyl-CoA carboxylase gene expression in porcine adipose tissue[J].Domestic Animal Endocrinology,1994,11(1):125-132.
[3] JEUKENDRUP A E.Regulation of fat metabolism in skeletal muscle[J].Annals of the New York Academy of Sciences,2002,967:217-35.
[4] SMITH S,WITKOWSKI A,JOSHI A K.Structural and functional organization of the animal fatty acid synthase[J].Progress in Lipid Research,2003,42(4):289-317.
[5] 周顺伍.动物生物化学[M].3版.北京:中国农业出版社,1999.
[6] FISCHER P P,WABITSCH M,HOCHBERG Z.Endocrinology of adipose tissue-an update[J].Hormone and Metabolic Research,2007,39(5):314-321.
[7] ASSAF S,HAZARD D,PITEL F,et al.Cloning of cDNA encoding the nuclear form of chicken sterol response element binding protein-2(SREBP-2),chromosomal localization,and tissue expression of chicken SREBP-1C and-2 genes[J].Poultry Science,2003,82:54-61.
[8] KIM J B,PASHA S,MARGART W,et al.Nutritional and insulin regulation of fatty acid synthetase and leptin gene expression through ADD 1/SREBP1[J]Journal of Clinical Investigation,1998,101:1-9.
[9] GERBENSF,JANSEN A,VAN ERPANTON JM,et al.The adipocyte fatty acid-binding protein locus:characterization and association with intramuscular fat content in pigs[J].Mammalian Genome,1998b(9):1022-1026.
[10] GERBENS F,VAN ERP ANTON J M,HARDERS F L,et al.Effect of genetic variants of the heart fatty acid-binding protein gene on intramuscular fat and performance traits in pigs[J].Journal of Animal Science,1999,77:846-852.
[11] ZHANG X,ZHAO S M,GE C R,et al.Effects of dietary digestible energy levels on meat quality in Wujin pig[J].Chinese Journal of Animal Nutrition,2008,20(4):377-387.
[12] ZHANG X,ZHAO S M,GE C R,et al.Effects of dietary energy levels on growth performance and carcass composition of Wujin pigs[J].Chinese Journal of Animal Nutrition,2008,20(5):489-450.
[13] ZHAO S M,WANG J,SONG X L,et al.Impact of dietary protein on lipid metabolism related gene expression in porcine adipose tissue[J].Nutrition&Metabolism,2010(7):6-18.
[14] 张宏福,张子仪.动物营养参数与饲养标准:中国肉脂型生长肥育猪饲养标准[M].北京:中国农业出版社,1998.
[15] BIKKER P,VERSTEGEN M W,CAMPBELL R G.Performance and body composition of finishing gilts(45 to 85)as affected by energy intake and nutrition in carlier life[J].Journal of Animal Science,1996,74:817-826.
[16] QUINIOU N,DOURMAD J Y,NOBLET J.Effect of energy intake on the performance of different types of pig from 45 to 100 kg body weight[J].Animal Science,1996,63:277-288.
[17] QUINIOU N,NOBLET J.The Effect of energy supply on the contribution of lean tissue to total body protein mass in pigs slaughtered at 100 kg[J].Animal Science,1997,65:509-513.
[18] 刘作华,杨云飞,孔路军,等.日粮能量水平对生长育肥猪肌内脂肪含量以及脂肪酸合成酶和激素敏感脂酶 mRNA表达的影响[J].畜牧兽医学报,2007,38(9):934-941
[19] 魏彩虹.猪脂肪沉积调控酶的研究进展[J].甘肃畜牧兽医,1997,27(4):29-32.
[20] KATSURADA A,IRIANI N,FUKUDA H,et al.Effects of nutrients and hormones on transcriptional and post-transcriptional regulation of acetyl-CoA carboxylase in rat liver[J].European Journal of Biochemistry,1990,190(2):435-441.
[21] IRITANIN,FUKUDA E,INOGUCHIK.A possible role of Z protein in dietary control of hepatic triacylglycerol synthesis[J].Journal of Nutritional Science and Vitaminology,1980,26:271-277.
[22] MILDNER A M,CLARKE S D.Porcine fatty acid synthase:cloning of a complementary DNA,tissue distribution of its mRNA and suppression of expression by somatotropin and dietary protein[J].Journal of Nutrition,1991,121:900-907.
[23] DING S T,SCHINCKELl A P,WEBER T E,et al.Expression of porcine transcription factors and genes related to fatty acid metabolism in different tissues and genetic populations[J].Journal of Animal Science,2000,78:2127-2134.
[24] 熊文中,杨凤,周安国.猪重组生长激素对不同杂交肥育猪脂肪代谢调控的研究[J].畜牧兽医学报,2001,32(1):1-4.
[25] BORGSTROM B,BROCKMAN H L.Adipose tissue lipases[C]//BELFRAGE P,FREDRIKSON G,STRALFORS P,et al.Lipases.Amsterdam:Elsevier,1984:365-416.
[26] SEMENKOVICH C F.Regulation of fatty acid synthase(FAS)[J].Progress in Lipid Research,1997,36:43-53.
[27] STOECKMAN A K,TOWLE H C.The role of SREBP-1c in nutritional regulation of lipogenic en-zyme gene expression[J].The Journal of Biological Chemistry,2002,277:27029-27035.
[28] BROWN M S,Goldstein J L.A proteolytic pathway that controls the cholesterol content of membranes,cells,and blood[J].Proceedings of the National A-cademy of Sciences of the United States of America,1999,96:11041-11048.
[29] GERBENS F,KONING D J,HARDERSF L,et al.The effect of adipocyte and heart fatty acid-binding protein genes on intramuscular fat and backfat content in Meishan crossbred pigs[J].Journal of Animal Science,2000,78:552-559.
[30] GARDAN D,LOUVEAV I,GONDRET F,et al.Adipocyte-and heart-type fatty acid binding proteins are both expressed in subcutaneous and intramuscular porcine(Sus scrofa)adipocytes[J].Comparative Biochemistry and Physiology,2007,148:14-19.
[31] YU Y H,GINSBERG H N.The role of acyl-CoA:diacylglycerol cyltransferase(DGAT)in energy metabolism[J].Annals of Medicine,2004,36(4):252-261.
