饥饿胁迫对瓦氏黄颡鱼(Pelteobagrus vachelli)幼鱼肌肉营养成分及血液指标的影响
2011-02-09仇明封功能张启焕刘汉文齐志涛
仇明 封功能 张启焕 刘汉文 齐志涛
由于光照、温度、空间和营养等生态环境的易变性,以及人类活动不断扩大及程度的加深,导致鱼类生态环境趋于恶化,鱼类时常会面临食物短缺而受到饥饿胁迫[1]。即使在养殖条件下,由于养殖密度过大,个体间相互竞争,投饵不足,投喂技术不当等原因导致鱼类出现因食物短缺引起的饥饿胁迫[2]。饥饿胁迫引起动物代谢、贮存物质消耗、免疫防御能力等的改变[3]。有关饥饿胁迫对鱼类生理、生态学影响的研究有助于揭示鱼类适应饥饿胁迫的生理生态学对策,对渔业资源管理及水产养殖实践有着重要的意义。自20世纪50年代开始,饥饿胁迫对鱼类生物学特征和生理机制的影响逐渐受到关注,并成为鱼类营养生理学研究的热点之一。当前,对鱼类饥饿胁迫的研究主要集中在鱼类存活状况、组织学、形态学、生理生化指标及内源性消化酶活力变化等方面,但鱼类的血液学指标也是鱼类生理学重要的研究内容,也是了解其健康状态、营养水平等的主要参数。
瓦氏黄颡鱼(Pelteobagrus vachelli)是我国养殖量最大并具有代表性的黄颡鱼品种,体型大、生长快,广泛分布于黄河、长江、珠江及黑龙江流域。其肉质细嫩,味道鲜美,营养价值高,深受消费者喜爱,具有潜在的广阔养殖前景。本试验用瓦氏黄颡鱼幼鱼为对象,研究饥饿胁迫对黄颡鱼幼鱼血液指标、肝脏中糖原和蛋白质含量、肌肉营养成分的影响,探索幼鱼在适应饥饿胁迫的生理、生态学特点和变化规律,对鱼类自然资源的保护、苗种培育、水产养殖等方面的实际生产具有重要的指导意义。
1 材料与方法
1.1 试验材料与设计
试验用鱼为瓦氏黄颡鱼夏花鱼种,出苗45 d的幼鱼,购自江苏省泰兴市水产良种场。试验随机化设计,分为饥饿胁迫0(对照)、10、20、30、40、50 d共6组,每组3个重复。
购得试验用鱼后,用10 mg/kg高锰酸钾溶液消毒,暂养10 d,饲喂自制饲料,选择规格整齐、体格健壮,1.52 g左右的瓦氏黄颡鱼720尾,随机分组,放入到18个规格为100 cm×60 cm×80 cm的水族箱中,每箱40尾。采用微流水系统养鱼,试验用水为曝气后的自来水,24 h增氧,平均水温(23±2)℃,氨态氮低于0.4 mg/l,pH值6.8~8.0,溶解氧大于5 mg/l,每天18:10时排污一次,水质符合渔业水质标准。
1.2 血液指标的测定
在饥饿0(对照)、10、20、30、40、50 d时,每箱随机抽取3尾鱼,尾静脉采血1 ml,不加抗凝剂,低温静置4 h,吸取上层血清放入离心管中,4℃、3 000 r/min离心10 min,全血制备血清,冷冻-20℃保存,待测血液中血糖浓度、溶菌酶活性。另随机抽取3尾鱼断尾取血,制备抗凝血(肝素钠抗凝),待测血液中红细胞、白细胞数量和血红蛋白含量。用美国贝克曼Cx-4型自动生化分析仪测定血糖浓度、溶菌酶活性,用贝克曼库尔特HMX型血细胞分析仪测定红细胞、白细胞数量和血红蛋白含量。血液生理、生化指标在盐城市疾病预防控制中心测定。
1.3 肌肉成分及肝脏中糖原和蛋白含量的测定
将每组每箱3尾鱼肌肉分别充分混合后,采用烘箱干燥法(GB6435—1986)、干法灰化法(GB/T6438—1992)、凯氏定氮法(GB/T6432—1994)、索氏抽提法(GB5009.6—2003)测定其总水分、粗灰分、粗蛋白、粗脂肪含量。采用蒽酮法测定肝脏中的糖原含量。用考马斯亮蓝法测定肝脏中的蛋白质含量。
1.4 数据统计与分析
原始数据经Excel2007初步整理后,用统计分析软件SAS8.1进行单因素方差分析(One-Way ANOVY),采用LSD法进行多重比较,用平均值±标准误(X±S.E.)表示。
2 试验结果
2.1 饥饿胁迫对瓦氏黄颡鱼幼鱼血液指标影响(见表1)
表1 饥饿胁迫对黄颡鱼幼鱼血液指标的影响(n=3S.E.)

表1 饥饿胁迫对黄颡鱼幼鱼血液指标的影响(n=3S.E.)
注:同行数据肩标小写字母不同者表示差异显著(P<0.05),小写字母相同者表示差异不显著(P>0.05)。下表同。
项目血糖含量(mmol/l)红细胞数量(×1012个/l)血红蛋白含量(g/100ml)白细胞数量(×109个/l)溶菌酶活性(U/ml)0 4.16±0.08a 1.35±0.04a 6.20±0.03a 4.18±0.04c 400.25±9.18a 10 4.14±0.07a 1.30±0.03ab 6.16±0.02a 4.18±0.08c 397.25±21.88a饥饿天数(d)20 4.13±0.02a 1.24±0.02b 6.21±0.04a 4.32±0.03b 385.75±5.91ab 30 4.09±0.11a 1.26±0.05b 6.14±0.03a 4.29±0.10b 377.25±5.74bc 40 3.93±0.02b 1.16±0.02c 6.08±0.05ab 4.47±0.09a 372.00±9.42bc 50 3.79±0.03c 1.07±0.05d 5.98±0.05b 4.56±0.08a 361.50±8.96c
从表1看出,与对照组相比,随着饥饿时间的延长,血液中红细胞数量呈下降趋势,饥饿胁迫20 d时开始呈显著性下降,饥饿50 d时下降了20.74%;白细胞数量与红细胞的数量呈相反的变化趋势,随着饥饿时间的延长呈上升趋势,饥饿胁迫50 d时白细胞数量增加了9.09%;血红蛋白含量在饥饿初期呈先降低后升高再降低的变化趋势,饥饿胁迫50 d后,血红蛋白含量下降了3.55%;血糖在饥饿初期缓慢下降(P>0.05),后期呈显著下降,饥饿胁迫50 d后下降了8.89%;血清中溶菌酶活性呈缓慢下降,饥饿50 d后,溶菌酶活性下降的幅度为9.68%,达到显著水平(P<0.05)。
2.2 饥饿胁迫对瓦氏黄颡鱼幼鱼肝脏中糖原、蛋白质含量的影响(见图1)

图1 饥饿胁迫对黄颡鱼肝脏糖原、蛋白质含量的影响
从图1可以看出,在正常生态环境中黄颡鱼幼鱼肝脏中蛋白质含量在11.0%~12.0%左右,糖原含量为125.0~130.0 mg/g。在遇到饥饿胁迫时,这两种贮能物质都呈下降趋势。饥饿胁迫20 d左右时,肝脏中蛋白开始显著性下降(P<0.05),但饥饿到40 d后下降趋势减缓(P>0.05),而糖原含量在饥饿到30 d时才开始显著性下降(P<0.05)。
2.3 饥饿胁迫对瓦氏黄颡鱼幼鱼肌肉成分的影响(见表2)
由表2可知,随着饥饿胁迫时间的延长,肌肉中粗灰分、水分含量逐渐升高,饥饿50 d后,粗灰分含量增加了34.11%,水分含量增加了4.85%,都呈显著性增加(P<0.05)。在50 d的饥饿胁迫试验中,粗脂肪含量呈先急剧下降,在饥饿胁迫前30 d内下降了7.23%,30~50 d试验中,呈相对缓慢下降的趋势。粗蛋白含量随饥饿胁迫时间的延长,先缓慢下降(P>0.05),而后呈急剧降低(P<0.05),到饥饿50 d左右时,粗蛋白含量下降了22.79%。
3 讨论
3.1 饥饿胁迫对黄颡鱼幼鱼血液指标的影响
鱼类血液指标被广泛运用于评价鱼类的健康状况、营养状况及对环境的影响状况,是良好的生理、病理和毒理学指标[4]。许多研究发现,饥饿胁迫时鱼类红细胞数、血红蛋白有减少现象[5-7]。本试验结果表明,随着饥饿时间的延长,血液中血糖含量、红细胞数量、溶菌酶活性和血红蛋白含量都呈显著性下降,白细胞数量呈显著性增加(P<0.05)。分析原因,随着饥饿时间的延长,黄颡鱼幼鱼养分缺乏,维持正常生理活动消耗存贮的糖原及血糖导致血液中血糖浓度显著下降(P<0.05)。维持体内各组织细胞活动所需的能量匮乏,新陈代谢减缓,鱼体造血功能受到抑制,血液中红细胞更新减缓,部分红细胞破裂,红细胞和血红蛋白含量降低,血液运输O2和排出CO2功能降低,能量代谢水平降低。调节机体酸碱平衡能力下降,溶菌酶活性降低,免疫抗病机能下降,白细胞数量增加[7-9]。
表2 饥饿胁迫对黄颡鱼肌肉营养成分的影响(n=3S.E)(%)
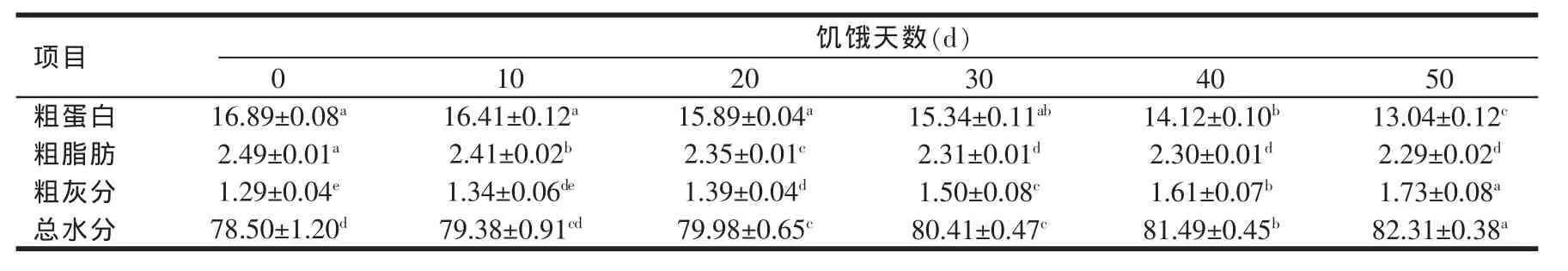
表2 饥饿胁迫对黄颡鱼肌肉营养成分的影响(n=3S.E)(%)
项目粗蛋白粗脂肪粗灰分总水分0 16.89±0.08a 2.49±0.01a 1.29±0.04e 78.50±1.20d 10 16.41±0.12a 2.41±0.02b 1.34±0.06de 79.38±0.91cd饥饿天数(d)20 15.89±0.04a 2.35±0.01c 1.39±0.04d 79.98±0.65c 30 15.34±0.11ab 2.31±0.01d 1.50±0.08c 80.41±0.47c 40 14.12±0.10b 2.30±0.01d 1.61±0.07b 81.49±0.45b 50 13.04±0.12c 2.29±0.02d 1.73±0.08a 82.31±0.38a
3.2 饥饿胁迫对黄颡鱼幼鱼内源性储存物质的影响
葡萄糖是生物体的必需燃料,葡萄糖通过血液输送到各个组织器官中,因而维持恒定的血糖浓度对鱼类正常生理活动有重要的作用。鱼类血糖的来源主要有食物中糖的消化吸收、肝糖原和肌糖原的分解、蛋白质及脂肪的糖异生作用等三条途径。这三条途径在激素及神经系统的不断调节下,使血糖浓度始终处于一种动态平衡,以维持鱼类正常的生理活动需要。在本试验中,黄颡鱼幼鱼不再从食物中得到糖类物质,因而血糖的维持主要靠贮存的糖原或糖异生作用,试验结果出现黄颡鱼幼鱼血糖浓度到饥饿40 d才显著性下降,而肝脏中蛋白质和糖类物质在饥饿20~30 d就开始显著性下降,肌肉中粗脂肪含量在饥饿胁迫10 d时呈显著性下降(P<0.05),肌肉中粗蛋白含量到40 d时显著性降低(P<0.05)。结果表明,瓦氏黄颡鱼幼鱼通过利用肝脏或肌肉中组成蛋白质的生糖氨基酸在转变为糖(葡萄糖),或肌肉中脂肪代谢转化为糖,补充消耗的血糖,维持血糖水平,为鱼类提供了生理活动的能量,当肌肉和肝脏中能源物质减少到一定程度后,血糖浓度也开始下降[10-11]。
研究表明[12-13],在饥饿期间,鱼类维持生理活动主要是通过动用身体贮存的能量和物质。因鱼的种类不同,动用哪种能源物质以及动用的先后顺序也有所不同。罗智等研究发现,饥饿胁迫前6周斑点叉尾鮰肌肉中蛋白质含量变化不显著(P>0.05),6周后呈显著降低(P<0.05)。随着饥饿时间的延长,糖原、脂肪及灰分含量呈显著降低(P<0.05),认为饥饿期间斑点叉尾鮰先通过利用肌肉中脂肪和糖原作为供能物质,在饥饿后期利用肌肉中的蛋白质作为能源物质[12]。但也有研究认为[13-14],一些鱼类在饥饿过程中首先利用蛋白质作为能源物质补充机体能量需要。这也许是因物种不同而不同。本研究发现,黄颡鱼幼鱼肌肉中粗脂肪在饥饿胁迫10 d后呈显著性下降(P<0.05),但饥饿到30 d以后下降趋势变缓,而粗蛋白含量随饥饿胁迫时间的延长先呈缓慢降低,饥饿40 d时呈急剧下降(P<0.05),这种下降的先后顺序表明黄颡鱼幼鱼在饥饿过程中主要是先动用肌肉中积累的脂肪,当脂肪消耗殆尽后开始消耗蛋白质作为身体的能量来源的,这与罗智研究结果相似。
[1] 章承军,刘健,陈锦辉,等.饥饿再投喂对缢蛏消化酶活力和抗氧化能力的影响[J].水产学报,2010,34(7):1106-1112.
[2] Kim M K,Lovell R T.Effect of restricted feeding regimens on compensatory weight gain and tissue changes in channel catfish Ictalurus punctatus in sponds[J].Aquaculture,1995,135(4):285-293.
[3] 薛明,柯才焕,王德祥,等.饥饿与再投喂对方斑东风螺生长、基本营养成分及RNA/DNA比值的影响[J].水产学报,2010,34(03):481-488.
[4] 董晓煜,张秀梅,张沛东.溶解氧与养殖密度对褐牙鲆幼鱼血细胞数量及血红蛋白含量影响的研究[J].海洋水产研究,2008,29(6):40-46.
[5] 胡一中,程宏毅,王鸿艳.饥饿对月鳢几项血液指标的影响[J].生物学杂志,2009,26(1):81-83.
[6] 杨成辉,蔡勋,刘霞,等.饥饿和再投喂对哲罗鱼幼鱼血液生理生化指标的影响[J].淡水渔业,2009,39(1):36-40.
[7] 乔志刚,张建平,牛景彦,等.饥饿和再投喂对鲇血液生理生化指标的影响[J].水生生物学报,2008,32(5):631-636.
[8] 程超,施光美.饥饿对鲫鱼血液生理生化指标和流变学性质的影响[J].中国农学通报,2008,24(1):516-519.
[9] 骆作勇,王雷,王宝杰,等.奥利亚罗非鱼饥饿后补偿生长对血液理化指标的影响[J].海洋科学进展,2007,23(3):340-346.
[10] 闫喜武,姚托,张跃环,等.冬季饥饿再投喂对菲律宾蛤仔生长、存活和生化组成的影响[J].应用生态学报,2009,20(12):3063-3069.
[11] Whyte J N C,Englaf J R,Carswell B L.Biochemical composition and energy reserves in Crassostrea gigas exposed to different levels of nutrition[J].Aquaculture,1990,90(2):157-172.
[12] 谭肖英,罗智,王为民,等.饥饿对小规格斑点叉尾鮰体重及鱼体生化组成的影响[J].水生生物学报,2009,39(01):39-45.
[13] 胡麟,吴天星.饥饿对鱼类生理生化的影响[J].水利渔业,2007,27(01):7-9.
[14] 张波,孙耀,唐启升.饥饿对真鲷生长及生化组成的影响[J].水产学报,2000,24(03):206-210.
