环杭州湾地区自然植被净初级生产力的变化特征及其成因
2011-01-09孙善磊周锁铨薛根元娄伟平吉宗伟石建红
孙善磊,周锁铨,薛根元,娄伟平,吉宗伟,石建红
(1.南京信息工程大学气象灾害省部共建教育部重点实验室,江苏南京210044;2.南京信息工程大学应用水文气象学院,江苏南京210044;3.宁波市气象局,浙江宁波315012;4.新昌县气象局,浙江新昌312500;5.绍兴市气象局,浙江绍兴313000;6.温州市气象局,浙江温州325027)
环杭州湾地区自然植被净初级生产力的变化特征及其成因
孙善磊1,2,周锁铨1,薛根元3,娄伟平4,吉宗伟5,石建红6
(1.南京信息工程大学气象灾害省部共建教育部重点实验室,江苏南京210044;2.南京信息工程大学应用水文气象学院,江苏南京210044;3.宁波市气象局,浙江宁波315012;4.新昌县气象局,浙江新昌312500;5.绍兴市气象局,浙江绍兴313000;6.温州市气象局,浙江温州325027)
利用1971—2006年环杭州湾地区25个气象站的降水、温度和云量资料及全球CO2年平均体积分数资料,采用LPJ全球动态植被模式(Lund-Potsdam-Jena Dynamic Global Vegetation Model),通过模拟环杭州湾地区的植被年净初级生产力(Annual Net Primary Productivity,ANPP),分析了该地区ANPP的变化特征,并探讨了植被ANPP变化的可能原因。结果表明:1)就环杭州湾地区,36 a间植被ANPP均表现出不同程度的增加,尤其以嘉兴市北部、绍兴市东部较明显;全区平均增加速率为1.524 3 g·m-2·a-2;2)通过多元线性回归分析发现,环杭州湾地区平均云量与植被ANPP的关系最为密切,偏相关系数为-0.517 5,而温度、降水与植被ANPP的关系不明显;同时,植被ANPP对气候变化的响应存在一定的地域性差异;3)在全区平均情况下,36 a间由温度下降、降水增加、云量减小、CO2体积分数升高引起的植被ANPP变化趋势分别为-0.081 3、-0.017 1、0.760 1、0.867 3 g·m-2·a-2,其对应的贡献率分别为-5.18%、-1.09%、48.38%、55.21%。由此可见,该地区植被ANPP变化的主要强迫因子是CO2体积分数和云量,而降水变化对植被ANNP的变化作用不大。
气候变化;植被净初级生产力;全球动态植被模式;LPJ模式;环杭州湾地区
0 引言
陆地生态系统与气候系统是相互作用和反馈的动态系统;而植被作为陆地生态系统的核心环节,在地表—大气之间能量、物质和动量交换中扮演着重要角色,是人类赖以生存与持续发展的生命支持系统(牛文元,1980;周广胜等,1997)。陆地植被净初级生产力(Net Primary Productivity,NPP)指的是绿色植物在单位时间和单位面积上所产生的有机干物质的总量(何勇等,2005)或碳的质量(本文NPP采用的单位),是植物自身生物学特性与外界环境因子相互作用的结果;作为表征植物活动的关键变量,反映出植被对大气中CO2固定的能力,是评价生态系统结构与功能特征和生物圈的人口承载力的重要指标。区域性乃至全球生物生产力及其空间分布,能使人类得以从宏观上作出如下估计:潜在的粮食资源的地理分布、人为提高区域性生产力水平的限度、不同国家和地区可能和现实的生产力水平(区域生态系统的最大容纳量)等。
近几十年,北半球气候变化显著(Serreze et al.,2000;Jones and Moberg,2003;蔡学湛等,2009;陈海山等,2009;周德刚等,2009;孙善磊等,2010a;姚洁等,2010),很大程度地改变了陆地生态系统,如碳平衡状况(Cao and Woodward,1998;Nemani et al.,2003),因此,开展气候变化下生态系统碳平衡的研究尤为重要。研究(Koca et al.,2006;Schaphoff et al.,2006)表明,生态系统碳平衡对气候变化的响应具有较大的空间异质性和不确定性,而生态系统尺度上的碳平衡模拟研究则有助于理解这种异质性和不确定性,有助于理解碳平衡的生态过程和驱动因素。
气候变化是世界各国关注的热点问题(IPCC,2007),随着由温室效应等引起的各种气候与环境问题的日益突出和国际气候谈判中对碳源、碳汇评价的客观需要,碳循环问题受到了人们的普遍关注(张家诚,1989;Keeling,1997;汪业勖和赵士洞,1998;Canadell and Mooney,1999;陶波等,2003;Gerber et al.,2004;赵玲和周锁铨,2005;Matthews et al.,2005;徐小锋等,2007;张佳华和姚凤梅,2007;Ju et al.,2007;Wang et al.,2007)。国内学者就气候变化对植被NPP的影响开展了很多有意义的工作。章基嘉等(1993)根据全国160个气象站资料,采用莱亨泊公式计算了光温生产潜力,给出了其时空变化特征及温度对光温生产潜力的影响。高素华(1994)从分析我国近40 a气温变化特征出发,初步分析了气温变化对农业、植被生产力的影响。刘文杰(2000)利用植物生理生态学特点及区域蒸散模式建立了NPP模型,探讨了NPP随气候变化的规律。曾慧卿等(2008)利用在我国有较好适用性的植被NPP模型,分析了近40 a气候变化对江西省自然植被净初级生产力的影响。周广胜和张新时(1995,1996)、周广胜等(1997)和郑元润等(1997)建立了联系植物生理生态特性和水热平衡关系的自然植被NPP模型,并利用该模型分析中国自然植被的NPP现状及全球变化后自然植被的NPP情况。
杭州湾位于我国东部沿海,地貌类型多样,属典型的亚热带季风气候;气候资源丰富,且气候地带性差异较大;而近几十年来有关该地区植被NPP空间分布、变化趋势以及其变化原因的研究很少见。另外,多数研究(倪健,2001;毛裕定等,2008)是以气候生产力模型为工具,对植被NPP的分布及变化特征进行研究;但是气候生产力模型自身存在一定的缺点(未考虑植被的生理过程和一些自然扰动,如火灾等),使得估算的气候生产力相对偏高。本文拟应用LPJ全球动态植被模式(Lund-Potsdam-Jena Dynamic Global Vegetation Model)模拟环杭州湾地区1971—2006年自然植被年净初级生产力(Annual Net Primary Productivity,ANPP),分析其变化特征;探讨植被ANPP对各因子的响应关系及各因子引起的植被ANPP的变化趋势和贡献率。这有利于认识气候变化和CO2体积分数升高对植被NPP的影响;并对合理开发、利用环杭州湾地区气候和土地资源,以及监测和预测该地区生态系统自然植被NPP的变化及应采取的策略提供科学依据。
1 资料、模型简介与研究方法
1.1 研究区域概况及气候背景
杭州湾为钱塘江口延伸的河口湾,是世界著名的喇叭型强潮河口湾。西起澉浦—西三闸断面,东至扬子角—镇海角连线。东西长90 km,湾口宽达100 km,自口外向口内渐狭,到澉浦仅为20 km。该地区邻近中国东南沿海,位于亚热带季风气候区,气候温和,空气湿润,雨水充足,光、温、水资源十分丰富,基本体现光温同步的特点。年平均温度为17℃,冬季盛行西北风,天气晴冷干燥;夏季多东南风,气温高,光照强,空气湿润;春秋两季气旋活动频繁,冷暖变化大。年平均降水量为1 600 mm,降水年内分布不均匀,春雨、梅雨、秋雨与伏旱、秋冬旱相互交替。该地区属于亚热带常绿阔叶林区域,自然植被为浙西山地丘陵青冈苦槠林分区。
1.2 数据资料及预处理
本文资料基于杭州市、嘉兴市、绍兴市和宁波市25个气象站的1971—2006年逐月降水、温度、云量数据;而1961—2006年全球CO2年平均体积分数数据来自LPJ工作组;土壤质地数据出自联合国粮农组织(Food and Agriculture Organization,FAO)、国际应用系统分析研究所(International Institute for Applied Systems Analysis,IIASA)和国际土壤资源信息中心(International Soil Reference and Information Centre,ISRIC)最新编制的全球土壤数据库。根据LPJ模式的要求,模式的输入数据为单站月气候平均值和CO2体积分数年平均数据;模式在每个气象站点上独立运行。研究区域以及气象站点分布见图1。
1.3 LPJ模式简介及适用性
LPJ全球植被动态模式为单点模式,它以站点经纬度、气候、土壤质地和大气CO2体积分数为输入变量,从植被动力学出发,以光合生物化学反应、冠层能量平衡以及光合作用产物在植物内部的分配规则和土壤水平衡为基础,考虑生态系统的自然死亡规律和自然干扰因素的影响,模拟生态系统的光合作用、呼吸作用、叶片形成和枝叶脱落、资源竞争、组织周转、种群建立及死亡、土壤微生物分解等过程,计算植物—土壤—大气之间碳和养分循环以及CO2和水的交换通量、光合作用强度和初级生产力以及植被和土壤之间的碳储量。模式以植物对环境的生态生理适应性和资源竞争力为基础模拟植被分布和组成。植被分10种植被功能型(plant function type,PFT;Hurtt,1998),包括8种树木植被(根据植被叶子物候、形状和地理纬度分布分类)和2种草本植被功能型(根据光合途径分类)。植物功能型的建立和死亡决定于一组基于20 a移动气候极值的生态生理限制条件,并考虑不同植物种类由于生态生理适应性、资源利用能力(如植物高度和根系深度)和光合效率的差异所产生的竞争优势或劣势。LPJ模式对植被组成结构变化、再分布和演替、动态变化、PFT种群间的竞争及土壤生物化学等都做了详尽描述。为了便于在网格中实现由植被个体到种群的尺度变换,LPJ模式的最小计算单位是PFT平均个体。对每种PFT都给定一套固定动力学和生态学参数,这样PFT种群就由一组描述平均个体状态的参数及其种群密度来确定。对树木植被,平均个体由其冠层面积、4个碳库(叶子质量、边材质量、心材质量和细根质量)确定,草本植被仅考虑叶子和细根的质量,并且设定其种群密度为1。
图1 环杭州湾气象站点分布Fig.1 The weather stations in the region around Hangzhou Bay
陆地生态系统模型是研究碳循环的重要方法,主要分为生物地理模型、生物地球化学模型和陆地生物物理模型3类(Pollard and Thompson,1995;Sellers et al.,1996;汪业勖和赵士洞,1998;Hurtt et al.,1998;毛留喜等,2006)。由其发展而来的全球动态植被模型,整合了以上三者的优点,可以模拟不同尺度生态系统结构和功能对气候变化响应的时间动态,因此其代表了全球碳循环模型的发展方向(毛留喜等,2006)。而植被动态模型LPJ作为全球动态植被模型的典型,对植物个体水平的模拟进行了细化,使植被组成、结构及其功能对气候变化响应的动态模拟过程更加精细(Smith et al.,2001;Sitch et al.,2003)。且LPJ作为更加优化的全球植被动态模型(Sitch et al.,2003;毛留喜等,2006),已经在全球不同区域得到了广泛应用(Hickler et al.,2004;Hely et al.,2006;田汉勤等,2007),尽管模型中尚未考虑一些生态过程(如氮循环),但诸多研究证明模型可以正确模拟生态系统结构和功能对气候变化的响应动态,例如森林FACE实验(Free Air CO2Enrichment Experiment)很好地验证了CO2体积分数增高情况下对NPP的模拟结果(Morales et al.,2007)。另一方面,由于所模拟的是普适性的生态过程,所以模型可以不经或少许地对参数校正后即可用于不同区域不同尺度的模拟研究(Gerten et al.,2005;Koca et al.,2006;田汉勤等,2007)。
1.4 研究方法及流程
首先,利用LPJ模式模拟出各站的自然植被的逐月NPP,计算出各站的ANPP;分析全区植被ANPP的变化趋势。然后,利用样条函数(spline function)的方法将植被ANPP插值到空间上,分析环杭州湾地区植被ANPP的36 a平均值及其变化趋势的空间分布。再后,利用多元线性回归分析的方法,分析植被ANPP对各因子的响应关系。最后,利用公式(1)计算各因子引起的植被ANPP变化趋势,并对其空间分布进行探讨。
根据LPJ模式的输入变量,自然植被ANPP(Yanpp)主要受降水(XP)、温度(XT)、云量(XCLD)、CO2体积分数(XCO2)的影响;而LPJ模式中辐射主要是利用云量来计算(Linacer,1968;Prentice et al.,1993;Haxeltine and Prentice,1996;Sitch et al.,2003;梁妙玲和谢正辉,2006),且大量研究表明净辐射与云量关系密切(Ohring and Clapp,1980;Shukla and Sud,1981;傅逸贤和彭莹辉,1988;Hartmann et al.,1992;Cawkwell and Bamber,2002;Nardinoand Georgiadis,2003;陈少勇等,2007;Lohmann et al.,2007;文小航等,2008);因此,本文利用ANPP与云量来表示其与辐射的关系。故可将植被ANPP的变化趋势分解为:
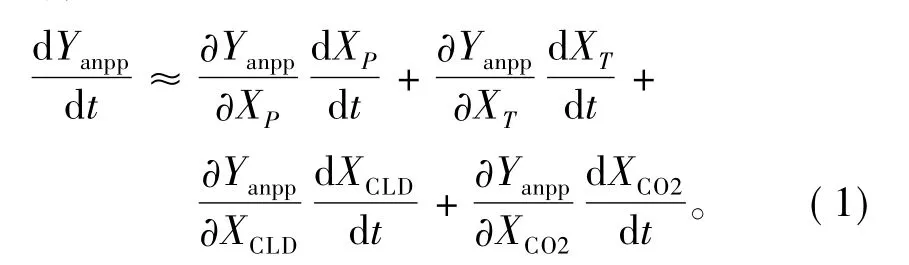
其中,式右第1—4项分别为降水、温度、云量和CO2体积分数变化引起的植被ANPP的变化趋势。
2 结果与分析
2.1 模拟结果比较
LPJ模式输出的环杭州湾地区的主要植被功能型有:温带针叶常绿林、温带阔叶常绿林、温带阔叶夏绿林及温带草场。经过实地考察,并结合中国植被类型分布图(侯学煜等,1982)比较发现:该地区主要以常绿阔叶林、常绿阔叶落叶混交林、阔叶落叶林、常绿针阔混交林为主,还有一些高、低草。因此,可以认为该模型能够较好地模拟出该地区植被类型。1971—2006年环杭州湾地区植被ANPP的36 a平均值介于518.3~615.0 g·m-2·a-1之间,与Wittaker and Likens(1973)、孙睿和朱启疆(2000)的研究结果基本吻合,略小于肖乾广等(1996)计算的最小阈值(主要是因为NOAA气象卫星资料的分辨率较低,由空间异质性引起),比张宪洲(1993)、孙善磊等(2010b)、毛裕定等(2008)的研究结果偏小,主要原因是Miami、Thornthwait、Chikugo等气候生产力模型没有考虑一些自然扰动因素(森林火灾等)的影响。
2.2 1971—2006年环杭州湾地区植被ANPP及其变化特征
由表1可见,1971—2006年嘉兴市、杭州市、绍兴市、宁波市和全区ANPP的36 a平均值分别为570.463 0、537.457 6、546.347 2、555.093 5和552.094 8 g·m-2·a-1。由图2a可以发现,在沿海地区,宁波市、嘉兴市ANPP随经度的减小而减小,绍兴市东北部ANPP存在一个相对大值区(570~585 g·m-2·a-1);在距沿海较远地区ANPP也呈现出经向分布,随经度的增加而减小。中部地区ANPP基本呈现出纬向分布特征,大概以绍兴市与杭州市的交界地区分别向南、向北逐渐递增。造成这种地区差异的主要原因是中部云量较多、温度较低、降水较少,而沿海及距沿海较远地区的相应条件优于中部地区,故ANPP较高。

表1 1971—2006年嘉兴市、杭州市、绍兴市、宁波市、全区不同年代的植被ANPP值及ANPP变化趋势Table 1 Natural vegetation ANPP in different periods and ANPP trend in Jiaxing,Hangzhou,Shaoxing,Ningbo and the region around Hangzhou Bay from 1971 to 2006
表1给出了嘉兴市、杭州市、绍兴市、宁波市、全区不同年代的ANPP值。可见,除嘉兴市ANPP最大值(589.164 8 g·m-2·a-1)出现在1991—2000年外,杭州市、绍兴市、宁波市、全区ANPP的最大值均出现在2001—2006年,分别为569.155 0、572.535 0、577.303 8、576.308 4 g·m-2·a-1。1971—2006年绍兴市的ANPP增长率最大,为2.080 8 g·m-2·a-2,宁波市的ANPP增长率最小,为1.000 0 g·m-2·a-2,全区平均增长率为1.524 3 g·m-2·a-2。
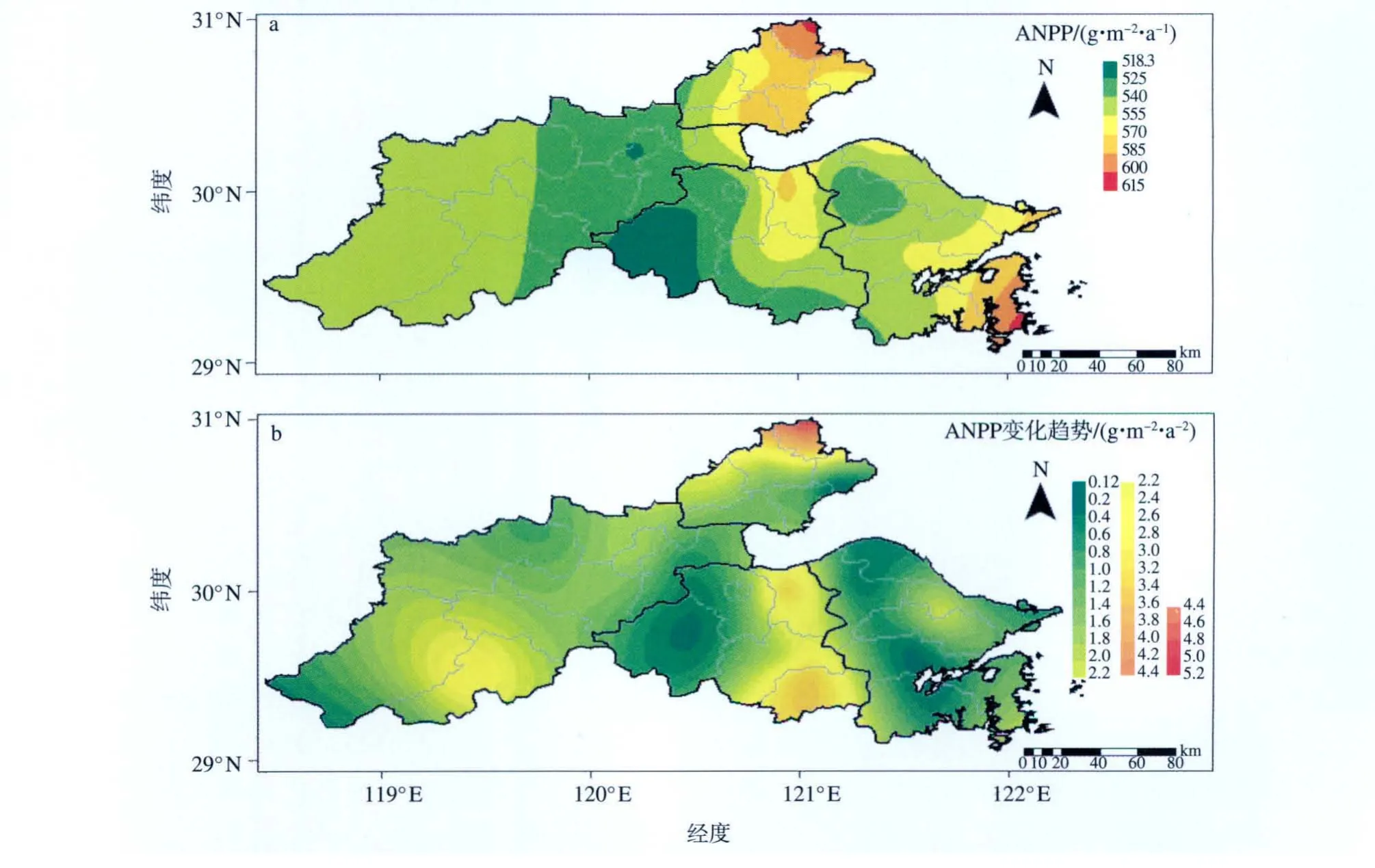
图2 1971—2006年环杭州湾地区植被ANPP的36 a平均值(a)及ANPP变化趋势(b)的空间分布Fig.2 Distributions of(a)36-yr average ANPP and(b)ANPP trend in the region around Hangzhou Bay from 1971 to 2006

图3 1971—2006年嘉兴市(a)、杭州市(b)、绍兴市(c)、宁波市(d)和全区(e)植被ANPP的时间变化曲线Fig.3 Interannual variation of regional average ANPP in(a)Jiaxing,(b)Hangzhou,(c)Shaoxing,(d)Ningbo,and(e)the region around Hangzhou Bay from 1971 to 2006
图3为1971—2006年嘉兴市、杭州市、绍兴市、宁波市、全区ANPP的时间变化曲线。可见,它们的年际变化曲线大体一致,都有略微上升的趋势;均在1992年出现了最大值;嘉兴市、杭州市、绍兴市、全区最低值出现在1975年,而宁波最低值出现在1976年。图2b为1971—2006年ANPP变化趋势的空间分布,可以发现,ANPP在全区都有不同程度的上升趋势,嘉兴市北部、绍兴市东部的上升趋势最为明显,杭州市南部地区上升趋势相对明显,而嘉兴市东部偏北、绍兴市东南部、宁波市西部的上升趋势相对较小。总体来看,随着气候变化,环杭州湾地区的ANPP是逐渐增加的,但是增幅存在一定的地域性差异,造成这种增幅地域性差异的主要原因是云量、温度、降水随时间的变化具有一定的地域性差异。
2.3 1971—2006年间各因子与环杭州湾地区自然植被ANPP多元线性回归分析
表2为1971—2006年环杭州湾地区4个影响因子的变化趋势。可以发现,全区、各市的温度均为上升趋势,且均通过了0.05信度的显著性检验;36 a间,全区温度上升趋势为0.043 7℃·a-1,其中,宁波市上升趋势最大(0.051 6℃·a-1),而绍兴市最小(0.036 6℃·a-1)。36 a间,全区降水以-0.721 7 mm·a-1的速率递减;除嘉兴市外,杭州市、绍兴市、宁波市均呈下降趋势,速率分别为-1.393 8、-1.298 0、-1.600 8 mm·a-1。全区、各市云量均呈现出不同程度的减小,其中嘉兴市、绍兴市减小趋势较显著,速率分别为-0.001 1 a-1、-0.000 9 a-1;全区云量变化趋势为-0.000 7 a-1。36 a间,CO2体积分数以1.569×10-6a-1的速率增加,且通过了0.001信度的显著性检验。
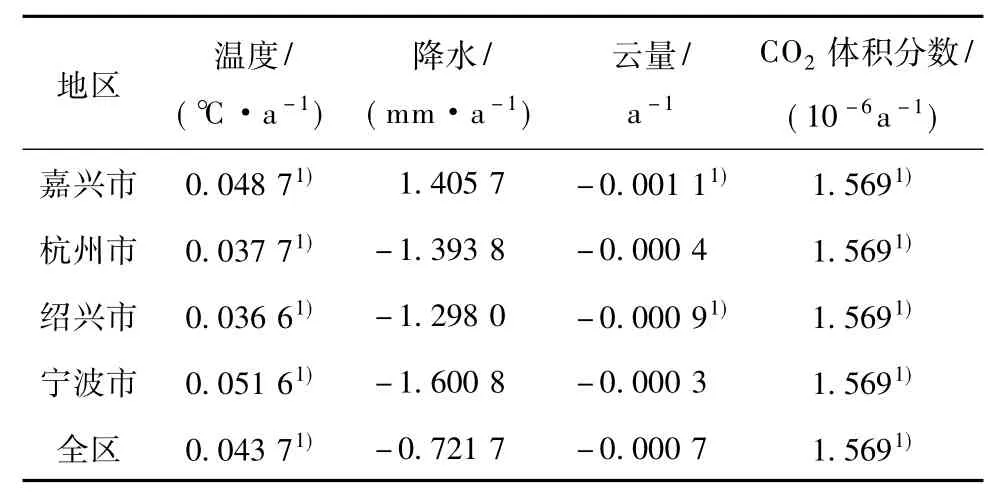
表2 1971—2006年4个影响因子的变化趋势Table 2 The trends of the four influencing factors from 1971 to 2006
本文拟采用多元回归分析的方法分析环杭州湾地区植被ANPP对温度、降水、云量、CO2体积分数的响应关系。表3给出了4个因子与植被ANPP的偏相关系数和复相关系数。可以看出,在4个因子中,4市、全区的云量与植被ANPP的偏相关关系最为密切,全区平均偏相关系数为-0.517 5,通过了0.05信度的显著性检验。不难理解,随着云量的逐渐减小,到达地面的太阳辐射增加,植被可以获取更多的光合有效辐射,有利于光合作用的进行,使得植被ANPP存贮增加。偏相关关系较差的为温度和降水,全区平均偏相关系数分别为-0.014 9、0.092 9;即随着温度的升高(下降),植被NPP有减少(上升)的趋势,而随着降水的增加(减少),植被NPP有增加(减少)的趋势。CO2体积分数与植被ANPP呈正的偏相关(0.120 7),即随着CO2体积分数的逐渐升高,植被更容易从大气中获取CO2进行光合作用,固定更多的碳,从而使得植被NPP增加。
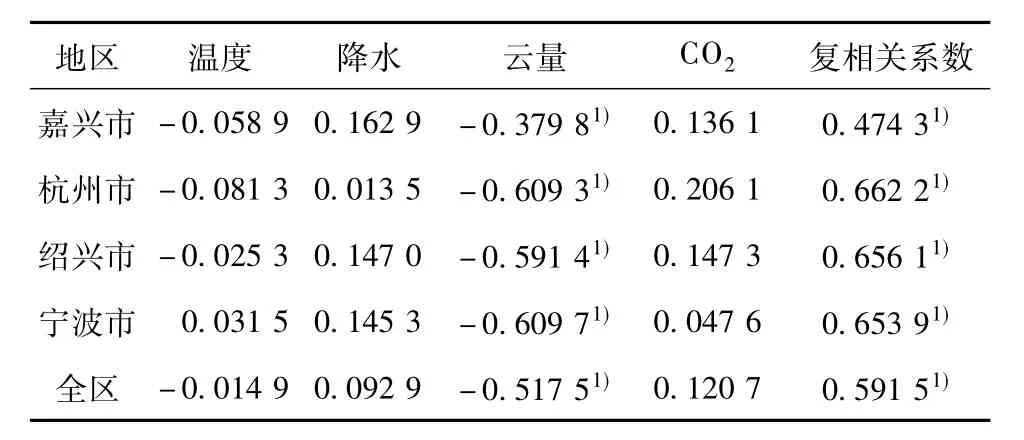
表3 4个因子与植被ANPP的偏相关系数及复相关系数Table 3 Partial correlation coefficients between the four factors and ANPP,and the multiple correlation coefficients
2.4 1971—2006年各因子对环杭州湾地区自然植被ANPP变化的贡献
表4给出了各因子变化引起的植被ANPP变化趋势及其贡献率。就全区平均而言,温度的升高、降水的减少都使得植被ANPP减小,而随着云量的减少及CO2体积分数的升高,植被ANPP增加(表3)。由温度变化、降水变化、云量变化和CO2体积分数变化引起的植被ANPP变化趋势分别为-0.081 3、-0.017 1、0.760 1和0.867 3 g·m-2·a-2,它们的贡献率分别为-5.18%、-1.09%、48.38%和55.21%。不难看出,各因子变化对植被ANPP变化贡献的主次关系为:CO2体积分数>云量>温度>降水。而在不同地区,由各因子变化对植被ANPP变化贡献的主次关系略有不同。
图4a为1971—2006年环杭州湾地区由降水变化引起的植被ANPP变化趋势的空间分布。可见,嘉兴市大部、杭州市西部及东部沿海地区、绍兴市北部沿海地区、宁波市西北沿海地区植被ANPP呈上升趋势,其中嘉兴市北部最明显,为0.2~0.4 g·m-2·a-2;杭州市东部、宁波市大部、绍兴市大部植被ANPP呈下降趋势,绍兴市南部地区有一负的低值中心。

表4 各因子变化引起的ANPP变化趋势及其贡献率Table 4 The trends of ANPP caused by four factors and their attribution rates
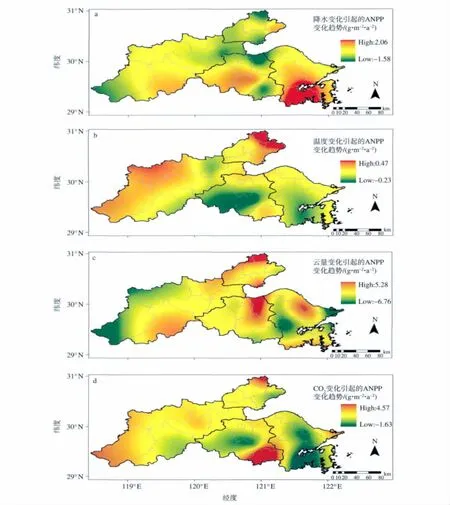
图4 1971—2006年环杭州湾地区由各因子变化引起的植被ANPP变化趋势的空间分布a.降水;b.温度;c.云量;d.CO2体积分数Fig.4 Distributions of ANPP trend caused by the four influencing factors in the region around Hangzhou Bay from 1971 to 2006a.precipitation;b.temperature;c.cloud;d.volume fraction of CO2
图4b为1971—2006年环杭州湾地区由温度变化引起的植被ANPP变化趋势的空间分布。可见,除了嘉兴市东北部、杭州市东南、绍兴市西南及中部、宁波市中部及南部为上升趋势外,环杭州湾大部分地区植被ANPP的变化呈下降趋势,绍兴市北部偏东、嘉兴市北偏西及南部偏东、杭州市北部偏东各有一负的低值中心。
图4c为1971—2006年环杭州湾地区由云量变化引起的植被ANPP变化趋势的空间分布。可见,嘉兴市东北部、杭州市东南部、绍兴市东北部、宁波市北部植被ANPP的上升趋势明显,杭州市西部、宁波市中部偏西及东北部下降趋势明显。
图4d为1971—2006年环杭州湾地区由CO2体积分数变化引起的植被ANPP变化趋势的空间分布。可见,大部分地区植被ANPP变化具有不同的上升趋势。然而,图4d中有3个负值中心,即由CO2体积分数变化引起的植被ANPP变化呈下降趋势。第一个负值中心位于嘉兴市西北部,范围较小,究其原因,可能是由于36 a间该地区云量呈上升趋势,光合有效辐射减少,使得植被固定的碳减少所致。第二、三个负值中心分别位于绍兴市中部和宁波市中南部,虽然CO2体积分数升高,但这两个地区温度升高、降水减小,气候条件有向“暖干”发展的趋势,不利于植被吸收CO2进行碳的固定,因而植被ANPP呈现下降趋势,这与胥晓(2004)的研究结果一致。
3 结论
利用1971—2006年环杭州湾地区25个常规气象站资料及全球CO2年平均体积分数数据,采用LPJ模式模拟了该地区植被ANPP,分析了其变化特征;并对植被ANPP对各影响因子的响应关系及各因子引起的植被ANPP变化趋势和贡献率进行了探讨,得到如下结论:
1)1971—2006年环杭州湾地区植被ANPP的36 a平均值为552.094 8 g·m-2·a-1;在2001—2006年最高,20世纪70年代最低;36 a间ANPP呈波状上升趋势,速率为1.524 3 g·m-2·a-2;全区ANPP都有不同程度的增加,但存在一定的地域性差异。
2)通过多元线性回归分析发现,4个影响因子中,云量与植被ANPP的偏相关关系最为密切,全区平均偏相关系数为-0.517 5,通过了0.05信度的显著性检验;由于水热条件的不同,这种响应关系也存在一定的地域性差异。
3)就全区平均而言,36 a间由各因子变化引起的植被ANPP变化的主次关系为:CO2体积分数(-0.867 3 g·m-2·a-2)>云量(0.760 1 g·m-2·a-2)>温度(-0.081 3 g·m-2·a-2)>降水(-0.017 1 g·m-2·a-2)。嘉兴市大部、杭州市西部及东部沿海地区、绍兴市北部沿海地区、宁波市西北沿海地区,由降水变化引起的植被ANPP变化呈上升趋势;而杭州市东部、宁波市和绍兴市大部,由降水变化引起的植被ANPP变化呈下降趋势。就大部分地区,由温度变化引起的植被ANPP变化呈下降趋势,且绍兴市北部偏东、嘉兴市北部偏西及南部偏东、杭州市北部偏东各有一负的低值中心;而嘉兴市东北部、杭州市东南、绍兴市西南及中部、宁波市中部及南部为上升趋势。在嘉兴市的东北部、杭州市东南部、绍兴市东北部、宁波市北部,由云量变化所引起的植被ANPP上升趋势明显;杭州市西部、宁波市中部偏西及东北部呈下降趋势。由CO2体积分数变化引起的植被ANPP变化趋势存在3个负值中心,分别位于嘉兴市西北部、绍兴市的中部和宁波市的中南部,而其他地区存在不同的上升趋势。
蔡学湛,温珍治,扬义文.2009.东亚夏季风异常大气环流遥相关及其对我国降水的影响[J].气象科学,29(1):46-51.
陈海山,范苏丹,张新华.2009.中国近50 a极端降水事件变化特征的季节性差异[J].大气科学学报,32(6):744-751.
陈少勇,董安祥,陈添宇,等.2007.祁连山总云量变化及其与气候变暖的关系[J].干旱区研究,24(1):98-102.
傅逸贤,彭莹辉.1988.云量在“地—气”系统净辐射中的作用[J].气象科学(4):69-75.
高素华.1994.气候变化对植物气候生产力的影响[J].气象,20(1):30-33.
何勇,董文杰,季劲均,等.2005.基于AVIM的中国陆地生态系统净初级生产力模拟[J].地球科学进展,20(3):345-349.
侯学煜,孙世洲,张经纬,等.1982.中国植被图[M].北京:中国地图出版社.
梁妙玲,谢正辉.2006.我国气候对植被分布和净初级生产力影响的数值模拟[J].气候与环境研究,11(5):582-592.
刘文杰.2000.西双版纳近40年气候变化对自然植被净第一性生产力的影响[J].山地学报,18(4):296-300.
毛留喜,孙艳玲,延晓冬.2006.陆地生态系统碳循环模型研究概述[J].应用生态学报,17(11):2189-2195.
毛裕定,苏高利,李发东,等.2008.气候变化对浙江省植物气候生产力的影响[J].中国生态农业学报,16(2):273-278.
倪健.2001.区域尺度的中国植物功能型与生物群区[J].植物学报,43(4):419-425.
牛文元.1980.作物最大生产力—以华北地区冬小麦为例[J].自然资源(3):44-56.
孙睿,朱启疆.2000.中国陆地植被净第一性生产力及季节变化研究[J].地理学报,50(1):36-45.
孙善磊,周锁铨,金博,等.2010a.淮海地区降水周期及突变特征分析[J].气象科学,30(2):221-227.
孙善磊,周锁铨,石建红,等.2010b.应用三种模型对浙江省植被净第一性生产力(NPP)的模拟与比较[J].中国农业气象,31(2):271-276.
陶波,李克让,邵雪梅,等.2003.中国陆地净初级生产力时空特征模拟[J].地理学报,58(3):372-380.
田汉勤,万师强,马克平.2007.全球变化生态学:全球变化与陆地生态系统[J].植物生态学报,31(2):173-174.
汪业勖,赵士洞.1998.陆地碳循环研究中的模型方法[J].应用生态学报,9(6):658-664.
文小航,尚可政,王式功,等.2008.1961—2000年中国太阳辐射区域特征的初步研究[J].中国沙漠,28(3):554-561.
肖乾广,陈维英,盛永伟.等.1996.用NOAA气象卫星的AVHRR遥感资料估算中国的净第一性生产力[J].植物学报,38(1):35-39.
胥晓.2004.四川植被净第一性生产力(NPP)对全球气候变化的响应[J].生态学杂志,23(6):19-24.
徐小锋,田汉勤.万师强.2007.气候变暖对陆地生态系统碳循环的影响[J].植物生态学报,31(2):175-188.
姚洁,陈海山,朱伟军.2010.北半球陆面过程对全球变暖响应特征的初步分析[J].大气科学学报,33(2):220-226.
曾慧卿,刘琪,殷剑敏.2008.近40年气候变化对江西自然植被净第一性生产力的影响[J].长江流域资源与环境,17(2):227-231.
章基嘉,徐祥德,苗俊峰.1993.气候变化对中国农业生产光温条件的影响[J].中国农业气象,14(2):11-16.
张家诚.1989.二氧化碳的气候效应与华北干旱问题[J].气象,15(3):20-25.
张佳华,姚凤梅.2007.陆面模式的植物叶光合生理的数值模拟试验及分析[J].气象科学,27(4):419-424.
张宪洲.1993.我国自然植被净第一性生产力的估算与分布[J].自然资源(1):15-21.
赵玲,周锁铨.2005.气候因子对寒温带落叶针叶林NPP影响的数值试验分析[J].气象,31(3):27-30.
郑元润,周广胜,张新时,等.1997.中国陆地生态系统对全球变化的敏感性研究[J].植物学报,39(9):837-840.
周德刚,黄荣辉,黄刚.2009.近几十年来长江上游流域气候和植被覆盖的变化[J].大气科学学报,32(3):377-385.
周广胜,张新时.1995.自然植被净第一性生产力模型初探[J].植物生态学报,19(3):193-200.
周广胜,张新时.1996.全球气候变化的中国自然植被的净第一性生产力研究[J].植物生态学报,20(1):11-19.
周广胜,张新时,高素华,等.1997.中国植被对全求变化反应的研究[J].植物学报,39(9):879-888.
Canadell J,Mooney H A.1999.Ecosystem metabolism and the global carbon cycle[J].Trends in Ecology and Evolution,14(6):249.
Cao M K,Woodward F I.1998.Dynamic responses of terrestrial ecosystem carbon cycling to global climate change[J].Nature,393:249-252.
Cawkwell F G L,Bamber J L.2002.The impact of cloud cover on the net radiation budget of the Greenland ice sheet[J].Annals of Glaciology,34:141-149.
Gerber S,Joos F,Prentice I C.2004.Sensitivity of a dynamic global vegetation model to climate and atmosphere CO2[J].Global Change Biology,10:1223-1239.
Gerten D,Lucht W,Schaphoff S,et al.2005.Hydrologic resilience of the terrestrial biosphere[J].Geophysical Research Letters,32,L21408.doi:10.1029/2005GL024247.
Hartmann D L,Ockert-Bell M E,Michelesen M L.1992.The effect of cloud type on earth’s energy balance:Global analysis[J].J Climate,5:1281-1304.
Haxeltine A,Prentice I C.1996.BIOME3:An equilibrium terrestrial biosphere model based on ecophysiological constraints,resource availability,and competition among plant functional types[J].Global Biogeochemical Cycles,10(4):693-709.
Hely C,Bremond L,Alleaume S,et al.2006.Sensitivity of African biomes to changes in the precipitation regime[J].Global Ecology and Biogeography,15(3):258-270.
Hickler T,Smith B,Sykes M T,et al.2004.Using a generalized vegetation model to simulate vegetation dynamics in northeastern USA[J].Ecology,85(2):519-530.
Hurtt G C,Moorcroft P R,Pacala S W,et al.1998.Terrestrial models and global change:Challenges for the future[J].Global Change Biology,4:581-590.
IPCC.2007.Climate Chang 2007:The Physical Science Basis[M].Cambridge:Cambridge University Press.
Jones P D,Moberg A.2003.Hemispheric and large-scale surface air temperature variations:An extensive revision and an update to 2001[J].J Climate,16:206-223.
Ju W M,Chen J M,Harvey D,et al.2007.Future carbon balance of China’s forests under climate change and increasing CO2[J].Journal of Environment Management,85(3):538-562.
Keeling C D.1997.Climate change and carbon dioxide:An introduction[J].National Academy of Science,94:8273-8274.
Koca D,Smith B,Sykes M T.2006.Modelling regional climate change effects on potential natural ecosystems in Sweden[J].Climatic Change,78:381-406.
Linacer E T.1968.Estimating the net-radiation flux[J].Agricultural Meteorology,5:49-63.
Lohmann U,Stier P,Hoose1 C,et al.2007.Cloud microphysics and aerosol indirect effects in the global climate model ECHAM5-HAM[J].Atmospheric Chemistry and Physics,7:3425-3446.
Matthews H D,Weaver A J,Meissner K J.2005.Terrestrial carbon cycle dynamics under recent and future climate change[J].J Climate,18:1609-1628.
Morales P,Hickler T,Rowell D P,et al.2007.Changes in European ecosystem productivity and carbon balance driven by regional climate model output[J].Global Change Biology,13:108-122.
Nardino M,Georgiadis T.2003.Cloud type and cloud cover effects on the surface radiative balance at several Polar sites[J].Theoretical and Applied Climatology,74:203-215.
Nemani R R,Keeling C D,Hashimoto H,et al.2003.Climate-driven increases in global terrestrial net primary production from 1982 to 1999[J].Science,300:1560-1563.
Ohring G,Clapp P.1980.The effect of changes in cloud amount on the net radiation at the top of the atmosphere[J].J Atmos Sci,37:447-454.
Pollard D,Thompson S L.1995.Use of a Land-Surface-Transfer Scheme(LSX)in a global climate model—the response to doubling stomatal-resistance[J].Global and Planetary Change,10:129-161.
Prentice I C,Sykes M T,Cramer W.1993.A simulation model for the transient effects of climate change on forest landscapes[J].Ecological Modelling,65:51-70.
Schaphoff S,Lucht W,Gerten D,et al.2006.Terrestrial biosphere carbon storage under alternative climate projections[J].Climatic Change,74:97-122.
Sellers P J,Los S O,Tucker C J,et al.1996.A revised land surface parameterization(SiB2)for atmospheric GCMs.Part II:the generation of global fields of terrestrial biophysical parameters from satellite data[J].J Climate,9:706-737.
Serreze M C,Walsh J E,Chapin F S,et al.2000.Observational evidence of recent change in the northern high-latitude environment[J].Climatic Change,46:159-207.
Shukla J,Sud Y.1981.Effect of cloud-radiation feedback on the climate of a general circulation model[J].J Atmos Sci,38:2337-2353.
Sitch S,Smith B,Prentice I C,et al.2003.Evaluation of ecosystem dynamics,plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model[J].Global Change Biology,9:161-185.
Smith B,Prentice I C,Sykes M T.2001.Representation of vegetation dynamics in the modelling of terrestrial ecosystems:Comparing two contrasting approaches within European climate space[J].Global Ecology and Biogeography,10:621-637.
Wang S,Chen J M,Ju W M,et al.2007.Carbon sinks and sources in China forest during 1901—2001[J].Journal of Environment Management,85(3):524-537.
Wittaker R H,Likens G E.1973.Primary production:The biosphere and man[J].HumanEcology,1(4):357-369.doi:10.1007/BF01536732.
Characters of natural vegetation net primary productivity and their driving factors in the region around Hangzhou Bay
SUN Shan-lei1,2,ZHOU Suo-quan1,XUE Gen-yuan3,LOU Wei-ping4,JI Zong-wei5,SHI Jian-hong6
(1.Key Laboratory of Meteorological Disaster of Ministry of Education,NUIST,Nanjing 210044,China;2.School of Applied Hydrometeorology,NUIST,Nanjing 210044,China;3.Ningbo Meteorological Bureau,Ningbo 315012,China;4.Xinchang Meteorological Bureau,Xinchang 312500,China;5.Shaoxing Meteorological Bureau,Shaoxing 313000,China;6.Wenzhou Meteorological Bureau,Wenzhou 325027,China)
Based on data of the precipitation,temperature and cloud cover of 25 stations in Hangzhou Bay region and the global annual volume fraction of CO2from 1971 to 2006,the annual vegetation net primary productivity(ANPP)is simulated by the LPJ model(Lund-Potsdam-Jena dynamic global vegetation model).The temporal and spatial characters of the ANPP in the region are analyzed and the driving factors of ANPP are further investigated.Results show that:1)The average increasing rate of ANPP is 1.524 3 g·m-2·a-2in the whole region.However,ANPP spatially show different increasing ratesfrom 1971 to 2006,with the significant rate in the north of Jiaxing and the east of Shaoxing.2)The multiple linear regression analysis shows that there is a close relationship between cloud cover and ANPP with-0.517 5 partial correlation coefficient.However,the correlation between ANPP and temperature(precipitation)is inconspicuous.Response of ANPP to climate change shows a certain geographical distribution for the different hydro-thermal conditions.3)In the whole region around Hangzhou Bay,the average trends caused by decline of temperature,increase of precipitation,reduction of cloud cover and enhancement of volume fraction of CO2are-0.081 3,-0.017 1,0.760 1,and 0.867 3 g·m-2·a-2,and their attributions to the change of ANPP are-5.18%,-1.09%,48.38%and 55.21%,respectively.Therefore,the changes of volume fraction of CO2and cloud cover play important roles to affect the change of ANPP,however,change of precipitation has little action to the ANNP.
climate change;vegetation net primary productivity;dynamic global vegetation model;LPJ model;the region around Hangzhou Bay
Q148
A
1674-7097(2011)06-0672-11
2010-04-20;改回日期:2011-09-01
国家自然科学基金资助项目(40775061/D0507);浙江省科技厅项目(2004C33082);绍兴市科学技术局项目(A23-2003131)
孙善磊(1982—),男,江苏沛县人,博士生,研究方向为陆—气相互作用、生态水文、气候变化及其预测,ppsunsanlei@163.com.
孙善磊,周锁铨,薛根元,等.2011.环杭州湾地区自然植被净初级生产力的变化特征及其成因[J].大气科学学报,34(6):672-682.
Sun Shan-lei,Zhou Suo-quan,Xue Gen-yuan,et al.2011.Characters of natural vegetation net primary productivity and their driving factors in the region around Hangzhou Bay[J].Trans Atmos Sci,34(6):672-682.
(责任编辑:倪东鸿)
