The Origin and Diversification of Osteichthyans and Sarcopterygians:Rare Chinese Fossil Findings Advance Research on Key Issues of Evolution
2010-12-30YUXiaoboZHUMinandZHAOWenjin
YU Xiaobo ZHU Min* and ZHAO Wenjin
1 Key Laboratory of Evolutionary Systematics of Vertebrates, Institute of Vertebrate Paleontology and Paleoanthropology (IVPP), CAS, Beijing 100044, China
2 Department of Biological Sciences, Kean University, Union, New Jersey 07083, USA
Living organisms represent only 1% of all the biota that has ever existed on earth.All organisms, living or extinct,are related to each other by sharing common ancestors at different levels,like twigs and branches connected to each other at different nodes on the great tree of life.One major task for paleontologists and evolutionary biologists is to find out how the diverse groups of organisms arose and how they are related to each other, thereby reconstructing the history of life and understanding the pattern and process of evolution.
As organisms evolve and diverge from common ancestors, they fall into a hierarchical (or “set-withinset”) pattern of groupings on the family tree, with members of each new group united by a common ancestor and characterized by novel biological features.For instance,within vertebrates, gnathostomes (or jawed vertebrates) arose as a new group when they acquired jaws as novel features, which set them apart from jawless agnathans (lampreys,hagfishes and their relatives).Within gnathostomes, four major groups developed: the bony osteichthyans(including all bony fishes and landdwelling tetrapods) with 53,633 living species, the cartilaginous chondrichthyans (sharks, rays and chimaeras) with 970 living species,and the long-extinct placoderms and acanthodians.Within osteichthyans,the actinopterygian lineage (with 26,981 living species) includes sturgeons, gars, teleosts and their relatives, while the sarcopterygian lineage (with 26,742 living species)includes lungfishes, coelacanths(Latimeria), their extinct relatives as well as all land-dwelling tetrapods(amphibians, reptiles, birds, and mammals) (Fig.1).
The quest for their origins has fascinated the human mind since the dawn of history.For many years, scientists have puzzled over questions such as: When and how didgnathostomes arise? What would the common ancestor of all osteichthyans look like? How do osteichthyans re late to chondr i cht hyans,placoderms and acanthodians? How did osteichthyans diverge into the actinopterygian and sarcopterygian lineages? When and where did sarcopterygians arise and diverge?How did some sarcopterygian fishes eventually give rise to land-dwelling tetrapods, which include humans?These and related questions not only lie at the center of evolutionary research but also have broad implications for science in general.Because of recent fossil fish findings from China, the global study on these key areas of vertebrate evolution has made significant progress and is now at the threshold of an exciting breakthrough in the near future.
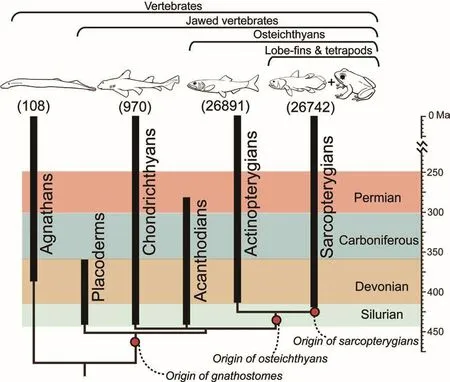
Fig.1 The family tree and geological time range of major vertebrate groups.Numbers of living species (in brackets) indicate the relative abundance and diversity of the four living groups (Nelson, 2006).
The most significant fossil fish finding from China is Guiyu, the oldest near-complete gnathostome fossil found from the 419-Myr-old muddy limestone in eastern Yunnan(Zhu et al., 2009; Fig.2).
Guiyu (“ghost fish”) gets its name because of its ghostly or bazaar combination of morphological characters.Within gnathostomes,Guiyu sides with osteichthyans by bearing derived macromeric scales,yet it resembles non-osteichthyan chondrichthyans, placoderms and acanthodians by having a primitive pectoral girdle and median fin spine.Within osteichthyans, Guiyu sides with sarcopterygians by having a two part braincase with a movable joint in the middle (plus other cranial features), yet it resembles early actinopterygians by having ornamented ganoine surface covering and by the shape of its cheek and operculo-gular bones.Computerized analysis of morphological characters places Guiyu as an osteichthyan in the basal segment of the sarcopterygian lineage (Zhu et al., 2009; Fig.3).Three other fossil fishes from Yunnan (Meemannia, Psarolepis and Achoania) cluster with Guiyu as basal or stem sarcopterygians, but they come from younger ages and consist of less complete materials when compared with Guiyu.The earliest geologic time of appearance, a nearcomplete preservation of whole body structures, and a unique combination of gnathostome vs non-gnathostome characters as well as sarcopterygian vs actinopterygian characters―all these make Guiyu highly significant for the global study on the origin of gnathostomes, the origin and diversification of osteichthyans,and the origin and diversification of sarcopterygians.

Fig.2 Reconstruction of “ghost fish” Guiyu as it may have lived in the Silurian waters 419 million years ago.Guiyu represents the earliest near-complete fossil gnathostome and fossil osteichthyan record.
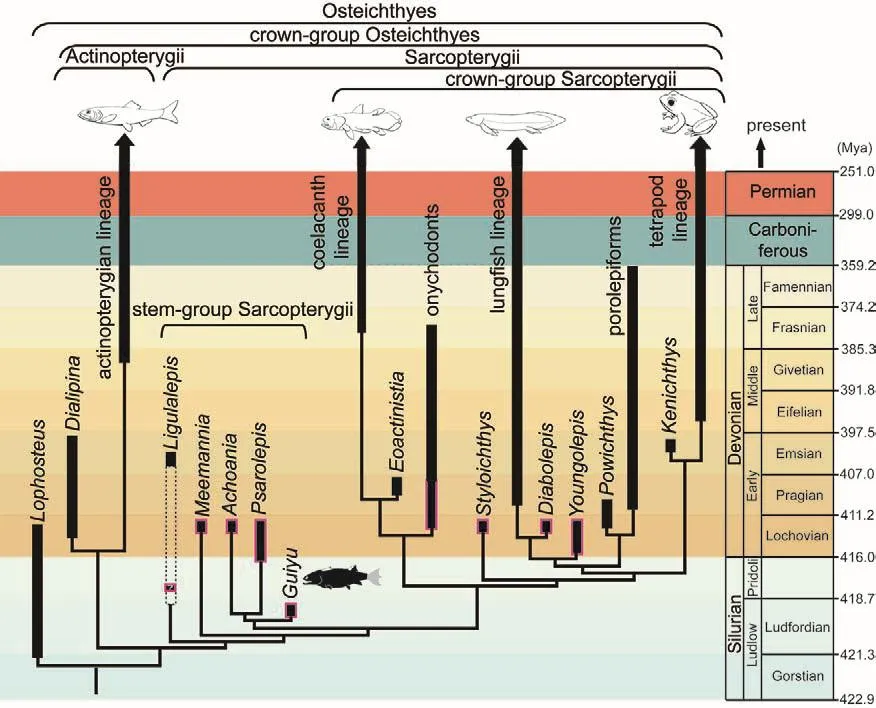
Fig.3 The family tree and geological time range of “ghost fish” Guiyu and other important early osteichthyans from China.Of the nine Late Silurian or earliest Devonian(Lochkovian) sarcopterygians, eight (framed in red) come from South China, suggesting that this region was once a center of origin and diversification for early sarcopterygians.
First, Guiyu provides critical fossil record to calibrate the molecular clock regarding the divergence time between two major osteichthyan lineages (i.e., actinopterygians and sarcopterygians).While modern biologists can determine the amount of molecular differences separating different living groups,this information can serve as a useful molecular clock only when it is calibrated by using fossils as specific historical time markers.Guiyu as a basal member of sarcopterygians has an accurate dating based on the Silurian conodont zonation (419 million years old), and this indicates that the minimal estimated time for the actinopterygian–sarcopterygian split must be at least 419 million years ago (instead of the previously 416-million-year-old estimate based on Psarolepis―another important early sarcopterygian fossil from Yunnan).With this new calibration,the molecular clock is now more accurate than before in gauging the time dimension for accumulated molecular changes between different living groups.
Second, Guiyu (together with seven other early sarcopterygians from the Silurian and earliest Devonian strata of southern China) establishes the paleobiogeographic importance of ancient South China region as a center of origin and diversification for sarcopterygians.So far, Yunnan has yielded eight out of the nine Late Silurian or earliest Devonian(Lochkovian) sarcopterygians,including four stem sarcopterygians(Guiyu, Meemannia, Psarolepis and Achoania) and four forms near the base of major subgroups of sarcopterygians (Styloichthys,Diabolepis, Youngolepis and an unnamed onychodont).During the Late Silurian–Lochkovian periods(about 411–422 million years ago), the South China region was a separate continent adjacent to eastern Gondwana, and its vertebrate fauna as a whole was highly endemic to the region, indicating little or no exchange between this region and other areas before the Middle Devonian period(397 million years ago).It was in this isolated or semi-isolated environment that major events in early sarcopterygian evolution unfolded and two subgroups developed, one giving rise to living lungfishes while the other giving rise to all tetrapods, including humans (Fig.3).
Third, Guiyu provides the most complete morphological information for establishing the morphotype (or primitive morphological model) of early gnathostomes, osteichthyans and sarcopterygians.Based on the morphotype for each major group, paleontologists can generate hypotheses about the sequential order of morphological changes from the origin of a group to specific evolutionary events in history, such as the branching of lineages or the conquering of new environments by new groups.
Gnathostomes may have first appeared in the Late Ordovician(more than 444 million years ago), but non-osteichthyan groups(chondrichthyans, placoderms and acanthodians) left little or no complete fossil specimens before the Early Devonian period.Similarly,previously known osteichthyan fossils from the Late Silurian are fragmentary and their lineage assignment is uncertain.Thus,Guiyu represents the earliest near-complete gnathostome and the earliest near-complete osteichthyan fossil known to science.Although other important osteichthyan fossils(Meemannia and Psarolepis from Yunnan, Ligulalepis from Australia,and Dialipina from the Canadian Arctic) have recently narrowed the morphological gap between different groups, they are mostly based on less complete materials and therefore are not as convincing as Guiyu in establishing the morphotype of early sarcopterygians and early osteichthyans.Guiyu provides a classical textbook case showing how scientific hypotheses can be tested through new discoveries.Before the discovery of Guiyu,the restoration of many features of Psarolepis (“speckled scale”)was sometimes regarded as weak inferences because of the isolated and disparate nature of fossil materials.The near-complete restoration of Guiyu (pending the discovery and description of complete Psarolepis specimen) provides the best possible authentication for most reconstructed features of Psarolepis (Yu, 1998;Zhu & Schultze, 1997; Zhu et al.,1999).Together with Psarolepis,Meemannia and other forms, Guiyu makes it possible to interpret features found in early members of both lineages (actinopterygians and sarcopterygians) in the common framework of deep osteichthyan phylogeny.Guiyu shows that the early sarcopterygians and actinopterygians retain widespread primitive osteichthyan and gnathostome features, such as the cheek and operculo-gular bone pattern and the anterodorsal process of the scale.
Lastly, Guiyu provides paleontologists with a new research agenda to focus more research resources and efforts on finding pre-Devonian osteichthyan fossils.Guiyu (and Psarolepis) suggest that early osteichthyans might carry median fin spine and spine-bearing pectoral girdles previously thought to exist only in non-osteichthyan groups.Consequently, many pre-Devonian spine-like materials can now be reexamined (and possibly reinterpreted) in the light of Guiyu and Psarolepis.
Meemannia (named in honor of Prof.Chang Meemann) is another fossil fish from Yunnan that has contributed significantly to the understanding of sarcopterygian evolution (Zhu et al., 2006).Typical or crown sarcopterygians are characterized by a special and now extinct tissue known as cosmine.Lack of similar tissue in living vertebrates and the previous lack of rudimentary forms of this tissue presented a major obstacle for scientists to understand its mode of origin and its function.Typical cosmine is found only in certain extinct sarcopterygians and consists of a pore-canal network embedded in mineralized dentine units and enamel.Like its mosaic morphological features, Meemannia reveals previously unknown histological features intermediate between the typical cosmine tissue (found in later sarcopterygians) and the ganoine-like tissues (found in actinopterygians and some acanthodians).Meemannia suggests a stepwise mode of origin for cosmine and lends support to the biological interpretation of the porecanal network as a vascular system supporting mineral deposition and resorption.Because of their sequential position on the early sectors of the sarcopterygian family tree, the cluster formed by Meemannia, Guiyu,Psarolepis, Achoania and Styloichthys(Fig.3) provides scientists with unprecedented record of character changes that helps to explain how sarcopterygians arose and eventually developed into the two later lineages,one leading to lungfishes, and the other leading to tetrapods, including humans.
In addition, these and other significant Chinese fossil fishes from Yunnan have become the“superstars” among paleontologists for many other reasons.For instance,Youngolepis (named after late Prof.Young Chungchien, pioneer of vertebrate paleontology in China) is an early occurring member (about 415 million years old) of the crown sarcopterygian group (Fig.3), and is one of the few osteichthyan fossils whose internal brain structures have been studied using 3-D reconstructions of serial grinding materials (Chang, 1982; Chang & Yu,1981).Kenichthys (named in honor of Prof.Ken Campbell of Australia)and Diabolepis (“devil scale”) are respectively the earliest members of the lineages leading to all tetrapods and leading to modern-day lungfishes(Chang & Yu, 1984; Chang &Zhu, 1993).Both Kenichthys and Diabolepis played a critical role in illuminating how tetrapods and lungfishes may have acquired their internal nostrils (or choana) as novel evolutionary features (Zhu& Ahlberg, 2004).Internal nostrils serve a vitally important function for all air-breathing animals (including humans), and its origin has puzzled paleontologists since the beginning of the 20thcentury.Styloichthys (“pillar fish”) shows what the common ancestor of modern-day lungfishes and tetrapods may have looked like and helps to explain the sequence in which the lungfish lineage and the tetrapod lineage acquired their respective characters (Zhu & Yu,2002).Achoania (“without choana or internal nostrils”) is closely related to Psarolepis and suggests that early or stem sarcopterygians may have had an eyestalk as found in sharks and rays (chondrichthyans) and extinct placoderms (Zhu et al., 2001).Last but not least, Qingmenodus represents one of the earliest known onychodonts (a subgroup of sarcopterygians distantly related to coelacanths) and helps to show how this ancient fish may have had the same feeding mechanism as the famous modern day living fossil Latimeria (Lu & Zhu, 2010).
The paleobiological roles of these fossil fish findings cannot be fully understood without detailed study of the associated faunas as a whole.Two significant vertebrate faunas have been established: the Late Silurian Xiaoxiang fauna and the Early Devonian Xitun fauna.In addition to Guiyu, Psarolepis and other (mostly microscopic or incomplete) remains of osteichthyans, the Xiaoxiang Vertebrate Fauna includes galeaspid agnathans, acanthodians, and diversified primitive placoderms.The research on the Xiaoxiang Vertebrate Fauna will significantly improve our understandings of early diversification of gnathostomes,especially when the morphological repertoire of acanthodians and the various primitive placoderms can be fully deciphered and analyzed in light of the new paradigm regarding the origin of osteichthyans from non-osteichthyan groups.The Early Devonian Xitun Vertebrate Fauna is characterized by the radiation and rich diversity of typical or crowngroup placoderms and crowngroup sarcopterygians (Zhao &Zhu, 2010).Given the exquisitely preserved fossils with fine-scaled stratigraphic control, study on the Xitun Vertebrate Fauna promises to yield productive results regarding the rapid diversification events responsible for the appearance of crown-group sarcopterygians.All this will provide a necessary framework to generate new hypotheses regarding the paleoenvironmental and paleogeographic aspects of this exciting chapter in the distant history of vertebrate evolution.
Main references
Chang, M.M., 1982.The braincase of Youngolepis, a Lower Devonian crossopterygian from Yunnan, south-western China.Stockholm: University of Stockholm.
Chang, M.M., & Yu, X.B., 1981.A new crossopterygian, Youngolepis praecursor, gen.et sp.nov., from lower Devonian of E.Yunnan,China.Scientia Sinica, 24: 89–97.
Chang, M.M., & Yu, X.B., 1984.Structure and phylogenetic significance of Diabolichthys speratus gen.et sp.nov., a new dipnoanlike form from the Lower Devonian of eastern Yunnan, China.Proceedings of the Linnean Society of New South Wales, 107: 171–184.
Chang, M.M., & Zhu, M., 1993.A new Middle Devonian osteolepidid from Qujing, Yunnan.Memoirs of the Association of Australasian Palaeontologists, 15: 183–198.
Lu, J., & Zhu, M., 2010.An onychodont fish (Osteichthyes, Sarcopterygii) from the Early Devonian of China, and the evolution of the Onychodontiformes.Proceedings of the Royal Society B, 277: 293–299.
Nelson, J.S., 2006.Fishes of the world, 4th edition.New Jersey: John Wiley & Sons, Inc.
Yu, X.B., 1998.A new porolepiform-like fish, Psarolepis romeri, gen.et sp.nov.(Sarcopterygii, Osteichthyes) from the Lower Devonian of Yunnan, China.Journal of Vertebrate Paleontology, 18: 261–274.
Zhao, W.J., & Zhu, M., 2010.Siluro-Devonian vertebrate biostratigraphy and biogeography of China.Palaeoworld, 19: 4–26.
Zhu, M., & Ahlberg, P.E., 2004.The origin of the internal nostril of tetrapods.Nature, 432: 94–97.
Zhu, M., & Schultze, H.-P., 1997.The oldest sarcopterygian fish.Lethaia, 30: 293–304.
Zhu, M., & Yu, X.B., 2002.A primitive fish close to the common ancestor of tetrapods and lungfish.Nature, 418: 767–770.
Zhu, M., Yu, X.B., & Ahlberg, P.E., 2001.A primitive sarcopterygian fish with an eyestalk.Nature, 410: 81–84.
Zhu, M., Yu, X.B., & Janvier, P., 1999.A primitive fossil fish sheds light on the origin of bony fishes.Nature, 397: 607–610.
Zhu, M., Yu, X.B., Wang, W., Zhao, W.J., & Jia, L.T., 2006.A primitive fish provides key characters bearing on deep osteichthyan phylogeny.Nature, 441: 77–80.
Zhu, M., Zhao, W.J., Jia, L.T., Lu, J., Qiao, T., & Qu, Q.M., 2009.The oldest articulated osteichthyan reveals mosaic gnathostome characters.Nature, 458: 469–474.
杂志排行

Bulletin of the Chinese Academy of Sciences的其它文章
- Dinosaurs of a Feather
- Amazing Reptile Fossils from the Marine Triassic of China
- Ongoing Research Projects at IVPP (A Selection)
- Advances in Vertebrate Paleontology and Paleoanthropology in China
- Recent Advances in Meso-Cenozoic Fish Research
- Dashankou Fauna: A Unique Window on the Early Evolution of Therapsids