小杜鹃对强脚树莺的巢寄生及其卵色模拟
2010-12-25杨灿朝
杨灿朝,蔡 燕,梁 伟
(1. 海南师范大学 生命科学学院,海口 571158;2. 中山大学 生命科学学院,广州 510275)
小杜鹃对强脚树莺的巢寄生及其卵色模拟
杨灿朝1,2,蔡 燕1,梁 伟1,*
(1. 海南师范大学 生命科学学院,海口 571158;2. 中山大学 生命科学学院,广州 510275)
于1999—2009年的鸟类繁殖季(4—8月)对贵州宽阔水自然保护区内的强脚树莺(Cettia fortipes)的繁殖进行监测,并采用光纤光谱仪,通过主成分分析、反射光谱、罗宾逊投射等方法对强脚树莺的卵色与小杜鹃(Cuculus poliocephalus)对其寄生的卵色模拟程度进行分析。研究结果表明,强脚树莺的繁殖成效在不同年份间均无显著差异,但其巢被捕食率和巢被寄生率都较高,分别为49.26%和9.18%。通过反射光谱分析,表明小杜鹃卵在色调和色度上高度模拟强脚树莺的卵,但其亮度高于宿主卵,而且在人眼无法探测的紫外光部分存在差异。
光纤光谱仪; 巢寄生;卵色模拟;寄生率;繁殖成效;小杜鹃;强脚树莺
杜鹃(Cuculus)产模拟寄生卵于寄主巢中,是因为在长期的共进化过程中寄主进化出高度的卵识别能力(Davies, 2000; Yang et al, 2010)。但是,巢寄生研究领域在国际上却长期存在一个似是而非的疑问,即有的寄主接受非模拟寄生卵;而有些被模拟寄生卵寄生的寄主却不具高度的卵识别能力。针对前者,Cherry & Bennett (2001)通过对非洲杜鹃和寄主卵色的光谱分析发现在人眼看来非模拟的杜鹃卵其实在紫外光上高度模拟寄主卵,在一定程度上解释了疑问,然而,对于后者一直没有很好的解释。强脚树莺(Cettia fortipes)产棕红色的卵,而小杜鹃(C. poliocephalus)在其巢中所产的寄生卵也为棕红色,其颜色在人眼看来非常相似,没有明显差异(Jiang et al, 2006)。在日本,小杜鹃寄生日本树莺(Cettia diphone),两者的卵色也均为棕红色。然而,被模拟卵寄生的日本树莺却不具高度卵识别能力(Higuchi, 1998),我们的初步研究也发现强脚树莺也不具备高度的卵识别能力。虽然小杜鹃所产的卵被认为是高度模拟的,但这都是建立在人类视觉的基础上的,并通过语言来进行描述,并没有对鸟类的卵色进行量化。如果以人类自定的标准来对颜色进行分类,可能会导致偏差,甚至错误的结论(Bennett et al, 1994),因为只有鸟类本身才是颜色信号的接收者(Endler, 1978, 1990)。绝大部分在日间活动的鸟类,其眼睛的圆锥细胞中至少存在 4种感光色素(Bowmaker et al, 1997),而且其对光谱最敏感的峰值处于紫外光区域,而这段区域对于人类来说是无法探测的(Chen et al, 1984)。所以,即使人类和鸟类一样能看到波长400—700 nm的光谱范围,但绝大多数鸟类只能探测到 400 nm以下的紫外光部分(Cuthill et al, 2000)。因此,许多在人眼看来是同样的颜色,实际上对于鸟类来说并不一定如此,反之亦然(Yang et al, 2009)。我们通过多年的研究,调查了小杜鹃对强脚树莺的巢寄生情况及其寄生率,并通过光纤光谱仪首次对小杜鹃与其寄主强脚树莺的卵色进行量化,分析杜鹃卵对宿主的模拟程度,探讨小杜鹃与强脚树莺在卵色上的共进化情况。
1 研究地区
贵州宽阔水自然保护区位于贵州绥阳县北部,东与正安县相邻,西与桐梓县接壤,为三县交界地区。总面积 26 231 ha,地理位置为 28º06'25''—28º19'25'' N,107°02'23''—107°14'09'' E。海拔在650—1 762 m之间,其中最高峰为海拔1 762 m的太阳山。森林覆盖率 80%,植被为以亮叶水青冈(Fagus lucida)林为主的原生性常绿落叶阔叶林,以及亚热带常绿阔叶林和常绿落叶阔叶混交林。该保护区位于大娄山东南侧,受西南季风和东南季风的影响,年平均气温 11.7—15.2℃,年平均相对湿度超过82%,年降水量介于1 300—1 350 mm之间,集中于4—10月,占年降水量的80%以上(Yu et al,2004)。
2 研究方法
2.1 鸟巢监测
于1999—2009年的鸟类繁殖季(4—8月),作者对强脚树莺的巢进行系统搜索,搜索范围涵盖宽阔水自然保护区各种生境类型,包括原生林、次生林、灌木林、竹林、茶地、茅草地、土坎和居民点等。用德国卡尔蔡司(10×42 BT)双目望远镜观察强脚树莺的繁殖行为,对繁殖个体进行针对性追踪,以定位其巢址。对每个发现的树莺巢,记录其筑巢时间、产卵时间、窝卵数、卵色、孵卵时间、巢寄生情况、杜鹃雏鸟生长情况和繁殖结果等资料。当巢被发现时处于孵卵阶段,用悬浮测验(floating test)推测产卵期(Hays & Lecroy, 1971)。用电子游标卡尺测量强脚树莺和杜鹃卵的长和宽(最小刻度 0.01 mm),用电子秤为卵称重(精确到0.01 g),卵容量的计算利用公式[0.51×卵长×(卵宽2)](Hoyt, 1979)。对发现的被寄生巢,将杜鹃卵和树莺卵放在统一的灰色板上,用佳能数码相机 (Canon EOS 500D)对其进行拍照。根据鸟巢的繁殖结果,计算其(1)被寄生率(parasitism rate);(2)被捕食率(predation rate);(3)孵化成效(hatching success):孵化的卵占所有卵的比例;(4)繁殖成效(reproductive success):每窝平均出飞的雏鸟数;(5)出飞成效(fledging success):成功出飞的巢中平均出飞的雏鸟数;(6)营巢成效(nesting success):繁殖成功巢占所有巢的比例,每窝至少有1只雏鸟出飞的巢为成功巢(Pribil, 1998)。
2.2 卵色的测量与数据处理
用荷兰 Avantes公司生产的 Avaspec-2048 USB2型光纤光谱仪对卵色进行测量,配套一个Avalight-Hal-S卤钨灯光源,反射探头(FCR-7UV200-2-ME)和一个探头固定器(RPH-1)。可测量的波长范围 200—1 000 nm,测量间隔为0.594 nm。用探头固定器将反射探头固定以相对鸟卵表面90°和距离鸟卵5 mm的位置进行反射光谱的测量,曝光时间(integration time)设置为100 ms。用聚四氟乙烯材料制成的白色漫射板(WS-2)来对测量进行标准化,它能反射所有波长范围的98%的光线。在测量反射光谱时,每枚鸟卵根据其长径从尖端到钝端被划分为5个区域,每个区域随机取测量点3个(Yang et al, 2009)。用Ava-Soft7.0分析卵色反射光谱数据,所提取的光谱范围为 300—700 nm,其中包括紫外光部分(300—400 nm)、蓝色光部分(400—475 nm)、绿色光部分(475—550 nm)、黄色光部分(550—625 nm)和红色光部分(625—700 nm)。描述颜色的反射光谱,研究光谱反射或形状的区别,用主成分分析(Principal Components Analysis)来处理数据(Cuthill et al, 1999; Cherry &Bennett, 2001)。利用主成分分析对反射光谱数据进行减维,此方法能将大量的相关性变量减少至少量而具代表性的正交变量(Bennett et al, 1997;Langmore & Bennett, 1999)。第一主成分(PC1)能描述平均反射变量,特别是非彩色亮度(Cuthill et al,1999; Hunt et al, 1998)。因为光谱间变量的绝大部分由此亮度变量组成,第一主成分通常解释了自然光谱中超过 90%的总变量,第二和第三主成分(PC2和PC3)代表了光谱形状的变量,所以分别跟色调和饱和度有间接相关(Endler & Théry, 1996; Grill &Rush, 2000)。卵色的分布模式用Endler & Paul (2005)的罗宾逊投射(Robinson Project)方法,并通Matlab 7.0 (MathWorks, Inc.)来实现,它将颜色的分布投射在以鸟类四种视锥细胞U、S、M、L敏感度为基础的四面体上,模拟从鸟类的视觉角度来看颜色,能客观分析和比较颜色的分布模式。U、S、M 和 L分别代表对紫外光、短波光、中波光和长波光的视锥细胞敏感度。统计检验在SPSS 13.0上进行,用单样本K-S检验(one-sample Kolmogorov -Smirnov test)来分析数据的正态性。当数据的正态性条件满足时,用t-测验(t-test)或方差分析(one-way ANOVA)来对均值间进行比较;若不服从于正态分布,则采用非参数检验,即 Mann-Whitney U test和Kruskal-Wallis test。当统计结果为P<0.05时,认为差异显著;P<0.01认为差异极显著;P>0.05则差异不显著。除非特别说明,给出均值为 Mean±SD。所有检验均为双尾(2-tailed)。
3 结 果
3.1 繁殖结果与巢寄生
共找到强脚树莺巢85个,其中76.81%筑于茶地,7.25%筑于竹林,7.25%筑于灌木,2.90%筑于茅草,5.80%筑于其他草本植物。巢高度为(0.88±0.38) m (n=82)。巢为侧面开口,由枯草叶编织,内垫鸡毛,巢宽(8.50±1.50) cm,巢深(11.00±1.41)cm,巢杯宽(4.76±0.81) cm,巢杯深(5.93±1.57 )cm(n=33)。卵为棕红色,重(1.45±0.13) g。体积(1.49±0.09) cm3(n=37)。
小杜鹃的寄生卵也为棕红色,但明显比强脚树莺的卵大(表 1),重(2.59±0.320 g,体积(2.60±0.02)cm3(n=2)。强脚树莺的窝卵数为(3.58±0.66)个(n=65),为了避免假重复,对其繁殖结果的统计去除了样本量小于15的年份,结果如表2,强脚树莺被捕食率和被寄生率较高,分别为49.26%和9.18%,不同年间无显著差异(χ2=0.77,df=3,P=0.96;χ2=0.85,df=3,P=0.84)。据观察啮齿类是主要捕食者之一,包括食卵的鼠类和侵占鸟巢的巢鼠类,其中巢鼠(Micromys minutus)侵占鸟巢做为自己的繁殖场所,被侵占的鸟巢包括刚筑好的巢、产卵期和孵卵期的巢。强脚树莺的孵化成效和营巢成效分别为47.54%和 30.20%,不同年间无显著差异(χ2=0.95df=3,P=0.81;χ2=0.22,df=3,P=0.98)。繁殖成效和出飞成效分别为(1.50±1.61)和(3.10±0.64),不同年间无显著差异(F(3,62)=0.59,P=0.62;F(3,16)=2.59,P=0.09,ANOVA。

表 1 强脚树莺和小杜鹃的卵大小和重量Tab. 1 Egg measurement of Brownish-flanked Bush Warbler and Lesser Cuckoo

表 2 强脚树莺的繁殖参数统计Tab. 2 Parameters of nest fate in Brownish-flanked Bush Warbler
3.2 卵色光谱分析
对强脚树莺卵色的主成分分析表明,前三个主成分总共解释了卵色99.84%的光谱变量(图1),其中第一主成分(PC1)解释了 91.33%,它的曲线是平缓的,代表了卵的亮度;第二主成分(PC2)和第三主成分(PC3)占总变量的 8.52%,其中 PC2的曲线是曲折的,它代表了卵色的色调;PC3则代表卵色的饱和度(色度)。罗宾逊投射(Robinson Project)表明,强脚树莺的卵色分布集中靠近长波光部分,离散程度低。通过箱丝图比较标准化的色调、色度和亮度,可以看出色度在这三种参数中的变异最大,亮度变异最小,但有一个异数点(图2)。小杜鹃寄生强脚树莺的卵色在可见光上高度模拟,其反射光谱的波形与寄主卵非常相似(图3)。在紫外光部分,两者的波形也很相似(图4),但在靠近300 nm处存在明显差异。
4 讨 论
4.1 繁殖成效与卵色变异
强脚树莺大部分巢集中在人工种植的茶园(76.81%),茶地生境单一,可能会招致集中性的被捕食和寄生。杜鹃在寄生过程中需要对处于繁殖阶

图1 强脚树莺卵色前三个主成分的协同系数Fig. 1 Principal component coefficient of PC1, PC2 and PC3 in the egg color of Brownish-flanked Bush Warbler
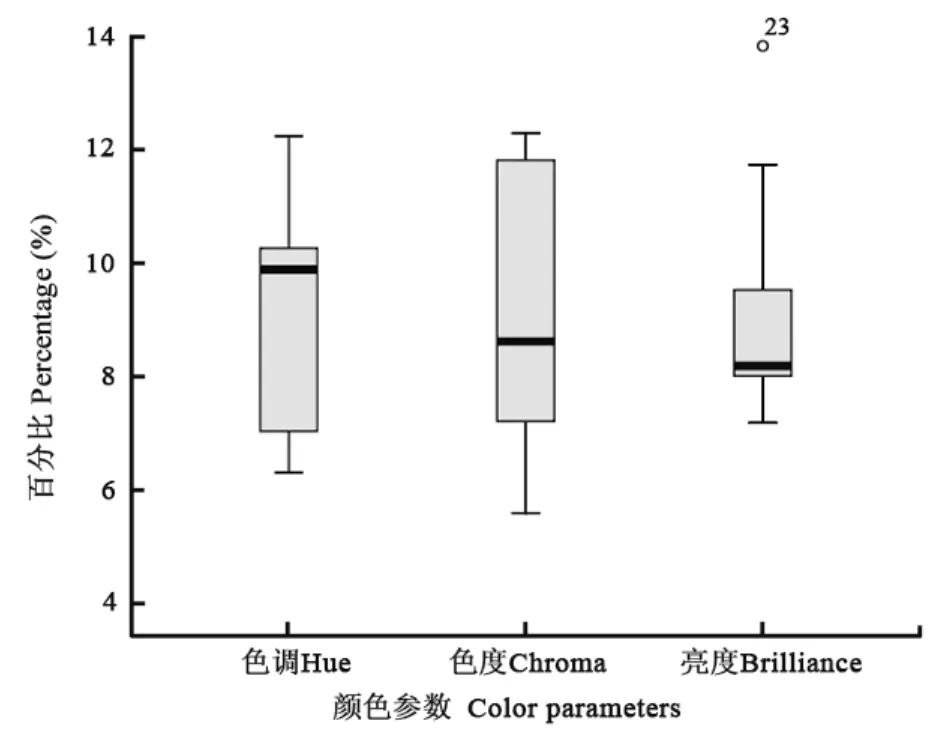
图2 强脚树莺卵色各颜色参数的标准化百分比Fig. 2 Standardized percentages of the egg color parameters in Brownish-flanked Bush Warbler
段的宿主进行暗中监视,宿主生境中的乔木等可以为杜鹃提供居高临下的监视点(Davies, 2000)。在我们的研究点,茶园面积虽大,但由许多斑块组成,有杉树坐落其中并有电线杆和电线贯穿茶园上方,为杜鹃提供了绝佳的监视点。据观察,经常有杜鹃雌鸟长时间停留在茶园中的杉树、电线杆或电线上“盯守”茶园,再者,这些监视点也可能有利于空中捕食者监视猎物,茶地的单一生境和理想的监视点可能是造成其被捕食率和被寄生率高的主要原因之一。较高的被捕食率和被寄生率导致强脚树莺的营巢成效和繁殖成效较低,但其孵化成效相对较高,这表明在雏鸟期也有相当一部分巢被捕食。
强脚树莺卵在标准化非彩色亮度上变异较小,但存在异数点(图2),这是由于强脚树莺卵壳颜色,偶尔会有色素明显变浅的情况,这往往出现在一窝卵的最后一枚卵,色素沉着变少使得卵色的亮度大大增高,这通过异数点表现出来。鸟类产到最后一枚卵的时候,由于腺体分泌的色素可能已耗尽(Nice,1937; Lowther, 1988),所以颜色变浅。Verbeek(1990)认为这是对捕食的适应,而Hockey (1982)则认为鸟巢中拥有不同颜色的卵能提高其隐蔽度,因为相比颜色很一致的一窝卵,前者更加难以被发现。Lloyd et al (2000)通过观察发现,色差异较大的巢似乎比较容易存活,然而,通过实验操作使得鸟巢卵色变得不一致并不能降低其被捕食率(Mason &Rothestein, 1987; Davies & Brooke, 1988)。有一个假说认为,因为鸟巢中的卵在鸟类产卵期的时候经常暴露在外,所以亲鸟需要投入更多的色素来隐藏其卵,而到了鸟类产最后一枚卵后,通常就进入孵卵期了,由于孵卵期亲鸟经常在巢中,卵很少暴露在外,所以最后一枚卵的色素浓度就显得不重要了,自然选择使得亲鸟将大部分的色素集中用于孵卵前的那几枚卵(Ruxton et al, 2001)。虽然没有直接的证据验证这个假说,但Martin et al.(2000)的研究发现,亲鸟较多的访巢会增加巢被捕食者发现的风险,这个结果与之前的假说相悖。

图3 寄生巢中小杜鹃与寄主强脚树莺的卵色可见光反射光谱Fig. 3 Visible egg reflectance of Lesser Cuckoo and Brownish-flanked Bush Warbler from parasitized nests

图4 寄生巢中小杜鹃与寄主强脚树莺的卵色紫外光反射光谱Fig. 4 Ultraviolet egg reflectance of Lesser Cuckoo and Brownish-flanked Bush Warbler from parasitized nests
4.2 杜鹃的卵色模拟
根据可见光反射光谱(图3),小杜鹃所产的寄生卵反射曲线波形高度模拟强脚树莺的卵,也就是在色调和色度上相似性高,但杜鹃卵的反射曲线高于强脚树莺的卵,这表明其卵的亮度高于寄主卵。在紫外光反射光谱上(图4),虽然小杜鹃卵和强脚树莺大体上波形一致,但在靠近300 nm处小杜鹃与寄主卵色的波形有明显不同。对于具有紫外感知视觉系统的脊椎动物,高能量的短波光对其上皮细胞,特别是视网膜组织具有光氧化效应(Collier &Zigman, 1987),对紫外波长的感知会导致视网膜光聚焦的不足,这是由于短波光具有强烈的非彩色色差和色散(Lythgoe, 1979),所以,对紫外光敏感的视觉系统带来相应的生理上的代价,相对于仅用人类可见光的视觉系统,其受到的自然选择强度是不一样的(Rick et al, 2006),也就是说,紫外光对于那些具有紫外感知能力的动物来说应该是具有重要生物学意义的。Cherry & Bennett (2001)对非洲的红胸杜鹃(Cuculus solitarius)和其众多寄主的卵色进行光谱分析表明,在人眼看来与寄主卵并不匹配的杜鹃卵,在紫外光部分跟寄主卵却是高度相似。Honza et al (2007)发现寄主可以利用紫外光和绿色光部分来识别自身卵和寄生卵。Avilés et al (2006)通过研究和总结认为鸟卵的紫外光可能对鸟类的卵色识别具有重要作用。综上所述,小杜鹃所产的寄生卵虽然在可见光上高度模拟,但在紫外光上有一定的差异,如果强脚树莺是利用紫外光来进行卵色识别,那么小杜鹃对其卵色的模拟就未能达到高度模拟的状态,这也就解释了为什么小杜鹃的寄主树莺不具有高度的卵识别能力。值得做进一步的研究工作,来测试寄主强脚树莺是否根据紫外光对卵色进行识别。
致谢:野外工作得到贵州省林业厅和贵州宽阔水国家级自然保护区的大力支持和协助!
Avilés JM, Soler JJ, Pérez-Contreras T. 2006. Dark nests and egg colour in birds: a possible functional role of ultraviolet reflectance in egg detectability [J].Proc R Soc Lond B, 273: 2821-2829.
Bennett ATD, Cuthill IC, Norris KJ. 1994. Sexual selection and the mismeasure of color [J].Am Nat, 144: 848-860.
Bennett ATD, Cuthill IC, Partridge JC, Lunau K. 1997. Ultraviolet plumage colors predict mate preferences in starlings [J].Proc Natl Acad Sci USA, 94: 8618-8621.
Bowmaker JK, Heath LA, Wilkie SE, Hunt DM. 1997. Visual pigments and oil droplets from six classes of photoreceptor in the retinas of birds [J].Vision Res, 37: 2183-2194
Chen DM, Collins JS, Goldsmith TH. 1984. The ultraviolet receptors in bird retinas [J].Science, 225: 337-339.
Cherry MI, Bennett ATD. 2001. Egg colour matching in an African cuckoo,as revealed by ultraviolet-visible reflectance spectrophotometry [J].Proc R Soc Lond B, 268: 565-571.
Collier R., Zigman S. 1987. The grey squirrel lens protects the retina from near-UV radiation damage [J]Prog Clin Biol Res, 247: 571-585.
Cuthill IC, Bennett ATD, Partridge JC, Maier EJ. 1999. Plumage reflectance and the objective assessment of avian dichromatism [J].Am Nat, 153:183-200.
Cuthill IC, Partridge JC, Bennett ATD, Church SC, Hart NS, Hunt S. 2000.Ultraviolet vision in birds [J].Adv Stud Behav, 29: 159-214.
Davies NB, Brooke M de L. 1988. Cuckoos versus reed warblers:adaptation and counteradaptation [J].Anim Behav,36: 262-284.
Davies NB. 2000. Cuckoos, Cowbirds and other Cheats [M]. London:Poyser.
Endler JA, Théry M. 1996. Interacting effects of lek placement, display behavior, ambient light, and color patterns in three neotropical forest-dwelling birds [J].Am Nat, 148: 421-452.
Endler JA. 1978. A predator's view of animal color patterns [J].Evol Biol,11: 319-364.
Endler JA. 1990. On the measurement and classification of colour in studies of animal colour patterns [J].Biol J Linn Soc, 41: 315-352.
Endler JA, Paul WM. 2005. Comparing entire colour patterns as birds see them [J].Biol J Linn Soc, 86: 405-431.
Grill CP, Rush VN. 2000. Analysis spectral data: comparision and application of two techniques [J].Biol J Linn Soc, 69: 121-138.
Hays H, Lecroy M. 1971. Field criteria for determining incubation stage in eggs of the common tern [J].Wilson Bull,83: 425-429.
Higuchi H. 1998. Host use and egg color of Japanese Cuckoos[M]//Rothstein SI, Robinson SK. Parasitic Birds and Their Hosts:Studies in Coevolution. New York: Oxford University Press, 80-93.
Hockey PAR. 1982. Adaptiveness of nest site selection and egg coloration in the African Black OystercatcherHaematopus moquini[J].Behav Ecol Sociobiol, 11: 117-123.
Honza M, Polačiková L, Procházka P. 2007. Ultraviolet and green parts of the colour spectrum affect egg rejection in the song thrush (Turdusphilomelos) [J].Biol J Linn Soc, 92: 269-276.
Hoyt DF. 1979. Practical methods of estimating volume and fresh weight of bird eggs [J].Auk, 96: 73-77.
Hunt S, Bennett ATD, Cuthill IC, Griffiths R. 1998. Blue tits are ultraviolet tits [J].Proc R Soc Lond B, 265: 451-455.
Jiang YX, Liang W, Yang CC, Sun YH. 2006. Brood parasitism on bush warbler by lesser cuckoo [J].Chn J Zool, 41(5): 31. [蒋迎昕, 梁 伟,杨灿朝, 孙悦华. 2006. 小杜鹃在强脚树莺巢中寄生繁殖. 动物学杂志, 41(5): 31.]
Langmore NE, Bennett ATD. 1999. Strategic concealment of sexual identity in an estrildid finch [J].Proc R Soc Lond B, 266: 513-550.
Lloyd P, Plaganyi E, Lepage D, Little RM, Crowe TM. 2000. Nest-site selection, egg pigmentation and clutch predation in the gound-nesting Namaque SandgrousePterocles namaque[J].Ibis, 142: 123-131.
Lowther PE. 1988. Spotting pattern of the last laid egg of the house sparrow[J].J Field Ornithol, 59: 51-54.
Lythgoe JN. 1979. The Ecology of Vision [M]. Oxford: Clarendon Press.
Martin TE, Scott J, Menge C. 2000. Nest predation increases with parental activity: separating nest site and parental activity effects [J].Proc R Soc Lond B, 267: 2287-2293.
Mason P, Rothstein SI. 1987. Crypsis versus mimicry and the evolution of shiny cowbird eggs [J].Am Nat, 130: 161-167.
Nice MM. 1937. Studies in the life history of the song sparrow [J].Tran Linn Soc New York, 4: 1-246.
Pribil S. 1998. Reproductive success is a misleading indicator of nest-site preference in the Red-winged Blackbird [J].Can J Zool, 76:2227-2234.
Rick IP, Modarressie R, Bakker TCM. 2006. UV wavelengths affect female mate choice in three-spined sticklebacks [J].Anim Behav, 71: 307-313.Ruxton GD, Broom M, Colegrave N. 2001. Are unusualy colored eggs a signal to potential conspecific brood parasites? [J]Am Nat, 157:451-458.
Verbeek NAM. 1990. Differential predation on eggs in clutches of northwestern crows: the importance of egg color [J].Condor, 92:695-701.
Yang CC, Cai Y, Zhang SP, Liang W. 2009. Quantitative analysis of bird egg color by using fiber spectrophotometer [J].Chn J Ecol, 28(2): 346-349.[杨灿朝, 蔡 燕, 张淑萍, 梁 伟. 2009. 利用光纤光谱仪量化和分析鸟类的卵色. 生态学杂志, 28(2): 346-349.]
Yang CC, Liang W, Cai Y, Shi SH, Takasu F, M?ller AP, Antonov A, Fossøy F, Moksnes A, Røskaft E, Stokke BG. 2010. Coevolution in action:disruptive selection on egg colour in an avian brood parasite and its host [J].PLoS ONE, 5(5): e10816.
Yu LF, Xie SX, Wu TL. 2004. Scientific Survey of Kuankuoshui Nature Reserve [M]. Guiyang: Guizhou Science and Technology Press. [喻理飞, 谢双喜, 吴太伦. 2004. 宽阔水自然保护区综合科学考察集[M].贵阳: 贵州科技出版社.]
Brood parasitism and egg mimicry on Brownish-flanked Bush Warbler (Cettia fortipes) by Lesser Cuckoo (Cuculus poliocephalus)
YANG Can-Chao1,2, CAI Yan1, LIANG Wei1,*
(1.College of Life Sciences, Hainan Normal University, Haikou571158,China;2.School of Life Sciences, Sun Yat-sen University, Guangzhou510275,China)
Nest fate of Brownish-flanked Bush Warbler (Cettia fortipes) was conducted in breeding seasons from 1999 to 2009 in Kuankuoshui Natural Reserve, Guizhou province. Predation rate, parasitism rate, hatching success,nesting success and reproductive success were surveyed and egg color was quantified using spectrophotometer. Principal component analysis, reflectance spectrum and Robinson Project were used to analyze the egg color of bush warbler and egg mimicry of Lesser Cuckoo (Cuculus poliocephalus) in parasitized nests. Our results indicated that the Brownish-flanked Bush Warbler suffered from high predation rate and relatively high parasitism rate of 49.26% and 9.18%, respectively. Reflectance analysis showed that the hue and chroma of Lesser Cuckoo eggs were highly mimetic but the egg brightness and ultraviolet reflectance were different from the bush warbler.
Spectrophotometer; Brood parasitism; Egg mimicry; Parasitism rate; Breeding success; Brownishflanked Bush Warbler (Cettia fortipes); Lesser Cuckoo (Cuculus poliocephalus)
Q959.732;Q959.739;S865.3503
A
0254-5853-(2010)05-0555-06
2010-03-29;接受日期:2010-07-28
2009年度教育部新世纪优秀人才支持计划(NCET-10-0111)资助*
Corresponding author),E-mail: liangw@hainan.net
杨灿朝(1981-),男,博士后,主要从事鸟类生态学研究
猜你喜欢
杂志排行

Zoological Research的其它文章
- Rates and patterns of microsatellite mutations in common carp (Cyprinus carpio L.)
- Ethogram of Yangtze finless porpoise calves (Neophocaena phocaenoides asiaeorientalis)
- Embryonic development of the concave-eared torrent frog with its significance on taxonomy
- Seasonal variation and synchronization of sexual behaviors in free-ranging male Tibetan macaques (Macaca thibetana) at Huangshan, China
- Identification of zRAP55,a gene .preponderantly expressed in StagesⅠandⅡ oocytes of zebrafish
- Niche partitioning between sympatric rhesus macaques and Yunnan snub-nosed monkeys at Baimaxueshan Nature Reserve, China