松墨天牛和花绒坚甲种群空间格局的地统计学分析
2010-12-21陈向阳邹运鼎毕守东
陈向阳,邹运鼎,毕守东
(1.黄山学院 生命与环境科学学院,安徽 黄山 245041;2.安徽农业大学 林学与园林学院,安徽 合肥 230036)
松墨天牛和花绒坚甲种群空间格局的地统计学分析
陈向阳1,2,邹运鼎2,毕守东2
(1.黄山学院 生命与环境科学学院,安徽 黄山 245041;2.安徽农业大学 林学与园林学院,安徽 合肥 230036)
应用地统计学的原理和方法研究了黑松、马尾松、湿地松树林松墨天牛和花绒坚甲种群的空间结构和空间相关性,结果表明3种松树林中松墨天牛种群的半变异函数曲线皆为球型,其空间格局为聚集型,2005年黑松、马尾松、湿地松上的松墨天牛种群空间变程依次为116.06、33.85、115.76,2004年黑松、马尾松上的松墨天牛种群空间变程依次为36.48、38.02;2005年黑松、马尾松、湿地松上的花绒坚甲种群空间变程依次为92.36、46.50、63.37,2004年黑松、马尾松上的花绒坚甲种群空间变程依次为空间变程依次为80.00、40.23,花绒坚甲种群的拟合半变异函数曲线也表现为球型,呈聚集空间格局,与松墨天牛种群变程和分布格局基本上一致,后者对前者在空间上有较强的追随关系,表明花绒坚甲与松墨天牛有一定依存关系。
松墨天牛;花绒坚甲;空间格局;地统计学
空间格局是种群的重要属性之一,研究掌握空间格局可以揭示种群的空间结构、种群下结构及物种间的空间联系,它是抽样技术的理论基础。过去的研究方法从早期的频次分布法到以后的扩散型指数法、回归模型分析法及其改进形式,其共同点是依赖于样本频次分布或均值一方差关系及其相应的判定准则确定空间分布格局。但是,这些方法未能考虑样本的空间位置和方向,不能区别不同空间格局的差异,并受样方和抽样区域大小的影响。[1-6]因此,需要一种既考虑样本值,又考虑样本空间位置及样本间距离的方法来研究种群的空间格局。地统计学(geostatistics)和前述方法不同,它考虑了样点的位置方向和彼此间的距离,直接测定空间结构相关性和依赖性,可用于研究有一定随机性和结构性的各种变量和空间分布规律。[7-11]松材线虫病(Bursaphelonchus xylophilus steiner)是我国松树上的毁灭性病害,松墨天牛(Monochamus alternatus)是该病的主要传播媒介,[12-14]控制松墨天牛种群则可以有效地控制松材线虫病的蔓延。花绒坚甲(Dastarcus helophoroides)是松墨天牛的重要寄生性天敌之一。[15-18]本文应用地统计学原理和方法研究不同树种松树上松墨天牛及其天敌花绒坚甲的空间分布,以揭示其空间格局及其空间关系。
1 材料与方法
1.1 试验地概况
敬亭山林场位于我国安徽省宣州市西北部(30°69’N,118°E),森林面积1336.2hm2,其中松林面积985.2hm2,占73.7%。在松林中,马尾松(Pinus massoniana)面积占 601hm2,密度在1800株·hm-2;湿地松(P.elliotii)和火炬松(P.taeda)占384.2hm2,密度在 900株·hm-2;还有少量黑松(P.thunbergii lamb)。样地选择在海拔220-250m,主要树种为马尾松,杉木(Cunninghamia Lanceolata),黑松、湿地松、火炬松,树龄20-22年,平均树高6-7m,胸径约6-14cm, 郁闭度为 0.5-0.8,1998年松毒 蛾(Dasychira argentata)暴发成灾,造成约13.3hm2的松林大量死亡,由于未及时清除死树,使得松墨天牛种群数量迅速增长,据宣州市宣州区森防检疫站2001年6月调查松墨天牛危害面积已达266.7hm2,松林死亡率2.3%,平均虫口6.3头/株。
1.2 样树的诱木的设置
2003年6月在该林场马尾松、黑松纯林设置样树诱木,作为诱木的松树面积各为1.0hm2,2004年6月在该林场的马尾松、黑松、湿地松的纯林及3种树种的混交林,再次设置样树诱木,作为诱木的3种松树纯林和混林面积各为1.2hm2,每种树为0.4 hm2,样树诱木的行株距均为20m。其诱木处理方法是:在诱木基部距离地面30cm用柴刀“品”字形各砍3个刀口,刀口与树干呈300夹角,入木深度1cm,用广东省林业科学研究院森林保护研究所研制的PA-28松墨天牛诱木剂原药液,与水按1:3比例稀释,使用专用的稀释施药塑料瓶进行施药,至药液不流出刀口为止,共处理300棵树。
1.3 调查方法
2004年5月26-28日在样地中砍伐纯林中的马尾松、黑松分别选择6株诱木进行解剖,2005年5月25-29日在样地中砍伐纯林和混交林中的马尾松、黑松、湿地松3种树种分别选择8株诱木进行解剖,记载诱木胸径大小,解剖诱木按每1m为一个区段,从砍伐的诱木基部开始编号,分段逐株解剖调查记载松墨天牛的侵入孔数,死、活幼数,死、活蛹数,成虫数和羽化孔数及其花绒坚甲的幼虫数和茧数。
1.4 分析方法
根据区域化变量的理论,[19,20]在空间上,昆虫种群数量是区域化变量。因此,可用区域化变量理论和方法进行研究。本文通过计算松墨天牛和花绒坚甲种群的实验半变异函数、拟合半变异函数模型、分析半变异函数结构来描述其空间格局和空间相关关系。对于观察的数据系列z(xi),i=1,2,…,n,样本半变异函数值可用下式计算:
其中,N(h)是被h分割的数据对(xi+xi+h)的对数,z(xi)和z(xi+h)分别是点xi和xi+h处样本的测量值,h是分割两样点的距离。
半变异函数有3个重要参数,即基台值、变程和块金值。变程指在变异函数的值达到平衡时的空间隔距离。基台值指达到平衡时的变异函数值,块金值是指变异曲线被延伸至间隔间距为零时的截距。这3个参数可反映昆虫种群空间格局或空间相关类型,给出该空间相关的范围。几种常用的理论模型如球型、指数型、高斯型和线型都可拟合实验半变异函数。球型半变异函数说明所研究的种群是聚集分布。它的空间结构是当样点间隔距离达到变程之前时,样点的空间依赖性随样点间距离增大而逐渐降低。指数型与球型模型类似,但其基台值是渐近线。非水平直线型的半变异函数表明种群是中等程度的聚集分布。其空间依赖范围超过研究尺度。如果是随机分布,则R*(h)随距离无一定规律性变化,完全随机或均匀的数据,R*(h)呈水平直线或稍有斜率,块金值等于基台值,表明抽样尺度下没有空间相关性。
2 结果与分析
2.1 不同林分几种松树上的松墨天牛及其天敌花绒坚甲种群数量
将2005年5月25-29日在样地中砍伐纯林和混交林中3种树种分别选择8株诱木进行逐株解剖调查,2004年5月26-28日在样地中砍伐纯林中的马尾松、黑松分别选择6株诱木进行解剖,记载松墨天牛列于表1及其天敌花绒坚甲种群数量列于表2。为了简化计算工作,首先对不同林分松墨天牛及其天敌花绒坚甲种群数量差异显著性进行分析。
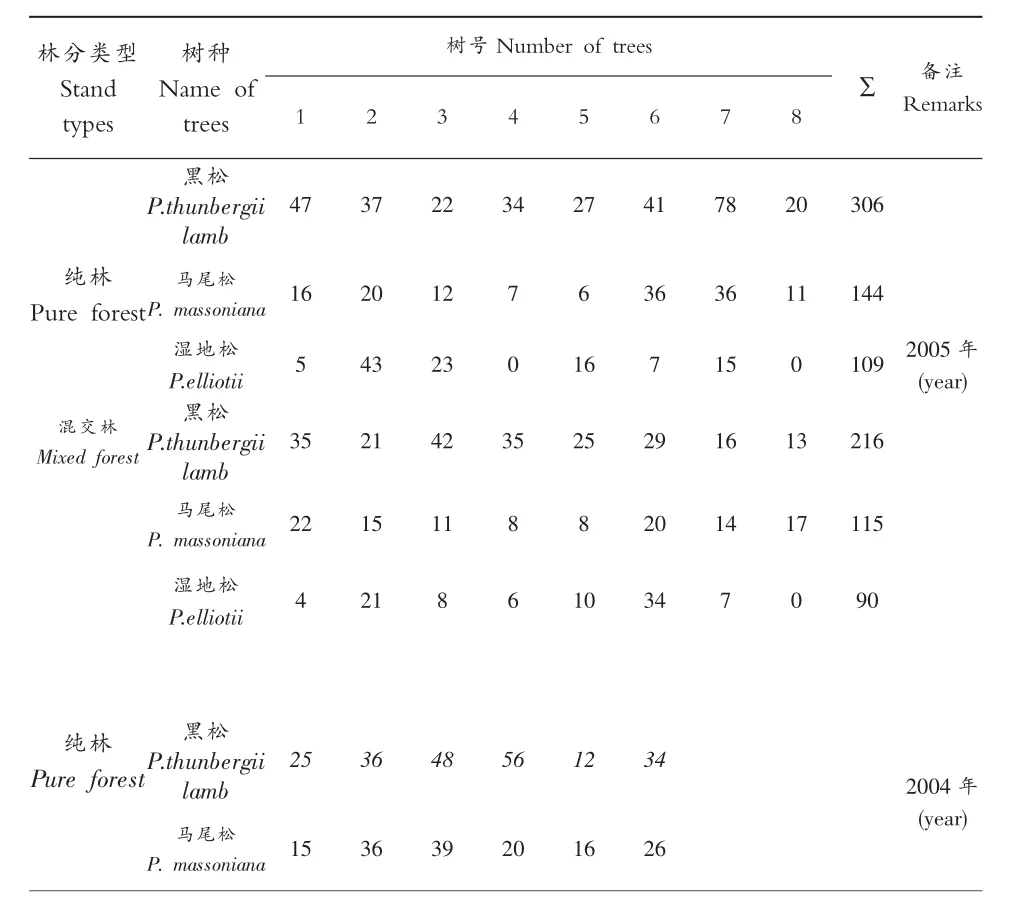
表1 不同林分不同松树松墨天牛种群数量(头/株)
将2005年调查的纯林和混交林同一种松树上的松墨天牛种群数量进行t检验,结果表明,黑松的t值为1.5069,马尾松的t值为0.2085,湿地松的t值为0.3708,当df=14时,t0.05=2.15,t值均小于2.15,同一种松树上的松墨天牛在纯林和混交林之间差异不显著,因此,两种林分松树上的松墨天牛可以合在一起进行地统计学分析。
同时对2005年调查的纯林和混交林同一种松树上的花绒坚甲种群数量进行t检验,结果表明,黑松的t值为1.257,马尾松的t值为0.6810,湿地松的t值为0.3489,当df=14时,t0.05=2.15,t值均小于2.15,同一种松树上的花绒坚甲种群数量在纯林和混交林之间差异不显著,因此,两种林分松树上的花绒坚甲种群数量可以合在一起进行地统计学分析。
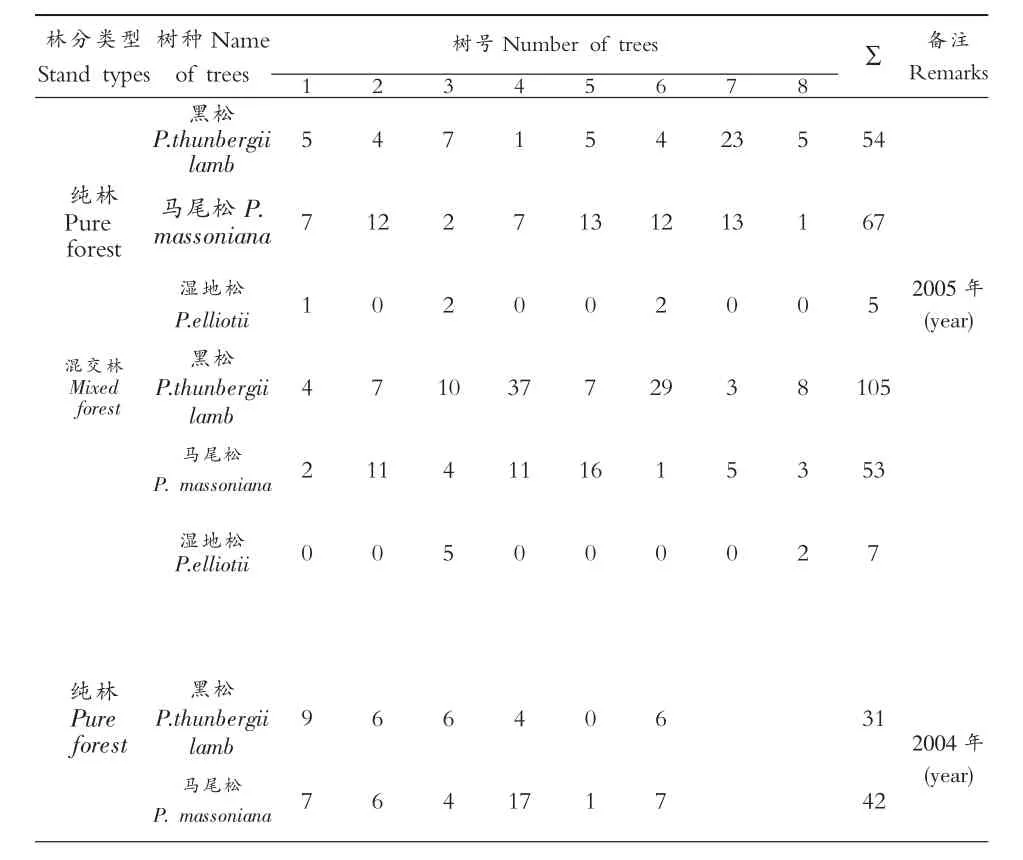
表2 不同林分不同松树花绒坚甲种群数量(头/株)
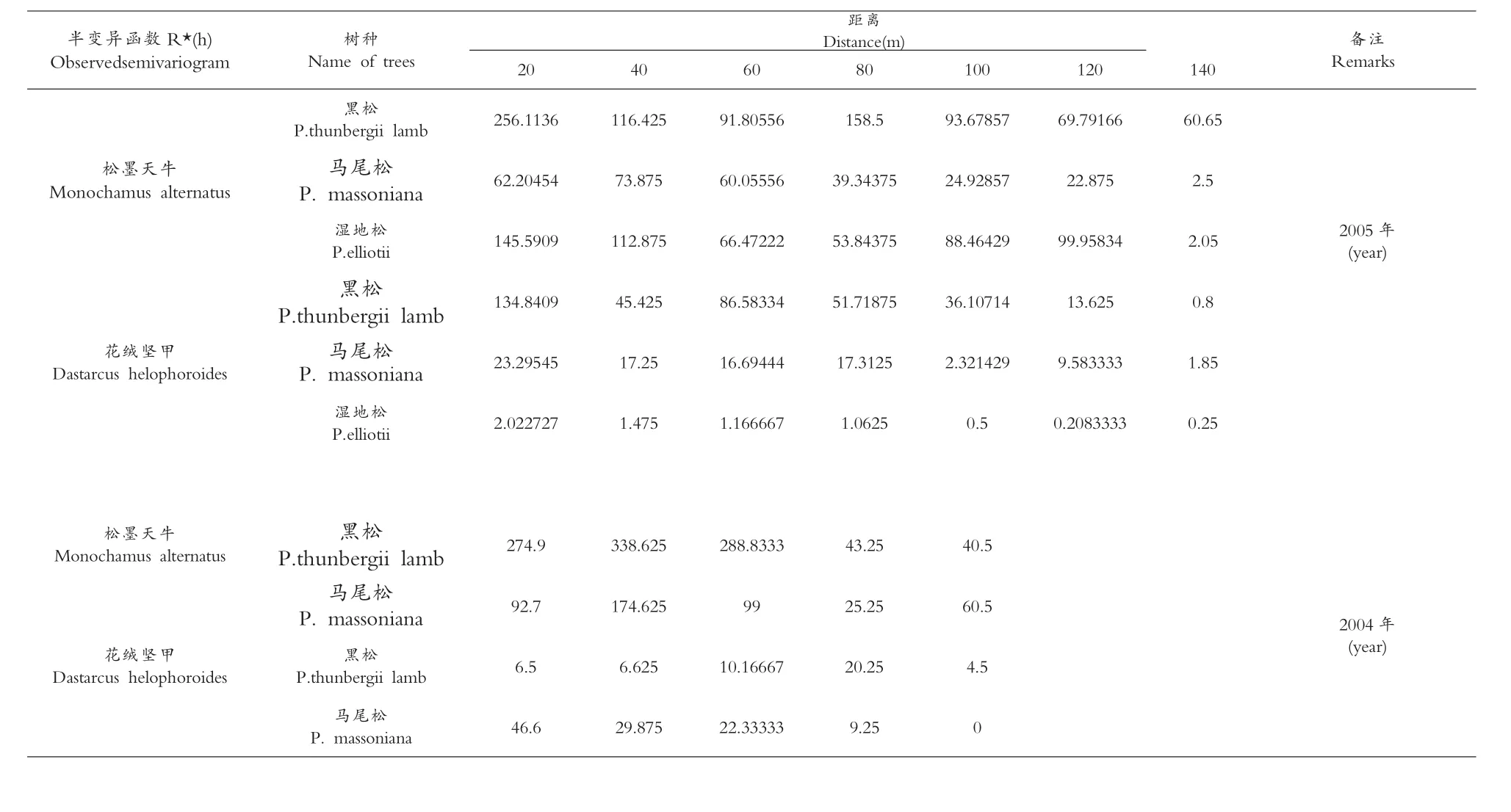
表3 不同树种上的松墨天牛、花绒坚甲种群半变异函数
2.2 松墨天牛种群数量的半变异函数和空间格局
将2004、2005两年的调查结果计算出不同树种的松墨天牛种群半变异函数值R*(h)列于表3,对松墨天牛种群空间结构进行分析,不同树种的松墨天牛种群半变异函数拟合模型均为球形,表明不同树种的松墨天牛种群的空间分布均为聚集分布,如表4所示。
松墨天牛种群在不同树种的半变异函数拟合模型具有较高的决定系数值,变动范围为0.7999-0.9968,表明实际半变异函数值与理论曲线的拟合度极高(图l)。不同树种松墨天牛种群半变异函数的变程RSD(m)即样点间的空间依赖性距离,在2005年黑松、马尾松、湿地松上的松墨天牛种群空间变程依次为116.06、33.85、115.76,在2004年黑松、马尾松上的松墨天牛种群空间变程依次为 36.48、38.02(见表4)。
2.3 花绒坚甲种群数量的半变异函数和空间格局
不同树种的花绒坚甲种群半变异函数值R*(h)列于表3,以此为实际值对理论半变异函数最优模型进行拟合(图l),不同树种的拟合半变异函数均为球形,说明不同树种的花绒坚甲种群分布为聚集分布(表3)。
花绒坚甲种群半变异拟合曲线的决定系数为0.7795-0.9937,表明半变异函数值与理论最优模型有极高的拟合度;不同树种的花绒坚甲种群半变异函数的变程RSD(m),即为样点间空间依赖性距离,在2005年黑松、马尾松、湿地松上的花绒坚甲种群空间变程依次为92.36、46.50、63.37,在2004年黑松、马尾松上的花绒坚甲种群空间变程依次为空间变程依次为80.00、40.24,如表4所示。
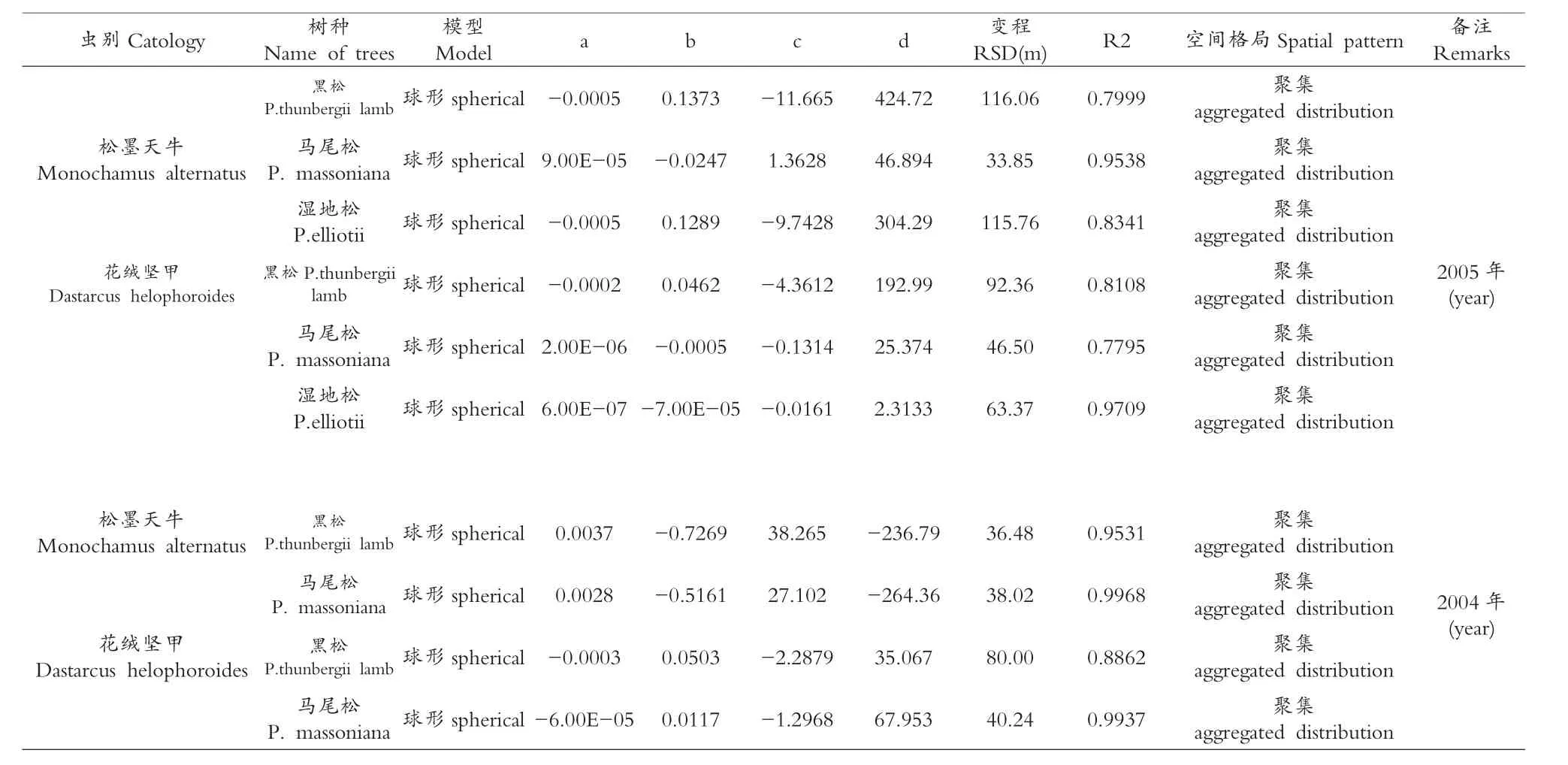
表4 不同树种上的松墨天牛、花绒坚甲种群的理论半变异函数模型参数
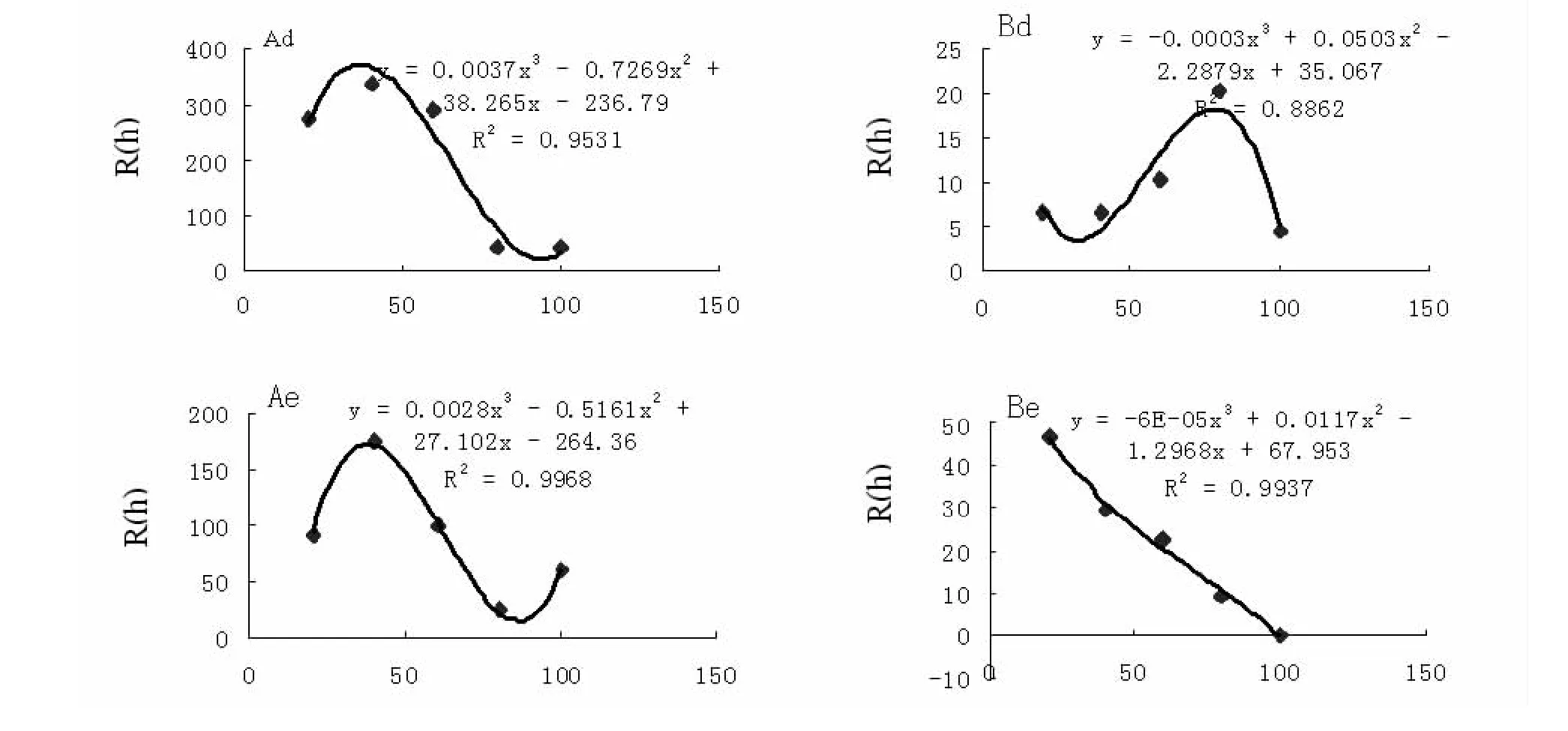
图1 松墨天牛(A)、花绒坚甲(B)的理论半变异函数曲线
3 讨 论
对松墨天牛及其天敌花绒坚甲种群数量在不同树种的地学统计学分析,得出不同树种的松墨天牛种群的空间格局均呈聚集分布,其半变异函数皆为球形曲线,样点间的个体是相互依赖的,其依赖程度随样点间的距离增大而逐渐降低,样点间的个体又是相互独立的。不同树种的花绒坚甲种群的空间格局均呈聚集分布,其半变异函数皆为球形曲线,不同树种的花绒坚甲种群变程与松墨天牛种群变程趋势基本一致。这充分说明了花绒坚甲种群对松墨天牛种群在空间位置上具有较强的追随关系。松墨天牛是蛀干性害虫,在松树诱木解剖中发现的几种天敌中,花绒坚甲的数量占绝对优势,表明了花绒坚甲是松墨天牛的优势种天敌。
[1]毕守东,邹运鼎,耿继光,等.棉蚜及龟纹瓢虫空间格局的地学统计学研究[J].应用生态学报,2000,11(3):421-424.
[2]邹运鼎,王弘法,巫厚长,等.龟纹瓢虫成虫对棉蚜的捕食作用[J].生物数学学报,1986,1(1):64-69.
[3]邹运鼎,毕守东,周夏芝,等.石榴园棉蚜及其天敌之间的关系[J].应用生态学报,2004,15(12):2325-2330.
[4]丁岩钦.昆虫种群数学生态学原理[M].北京:科学出版社,1980.
[5]刘树生,汪信庚,吴晓晶,等.甘蓝和白菜上桃蚜种群的空间格局及其时序动态[J].昆虫学报,1996,39(2):158-165.
[6]Sawyer AJ.Inconstancy of Talor’s b:simulated sampling with different quadrat sizes and spatial distributions[J]. Res Popul Ecol,1989,31:11-24.
[7]邹运鼎,毕守东,王祥胜,等.麦长管蚜及蚜茧蜂空间格局的地学统计学研究[J].应用生态学报,2001,12(6):887-891.
[8]黄寿山,胡慧建,梁广文,等.二化螟越冬空间分布图式的地理统计学分析[J].生态学报,1999,19(2):250-253.
[9]李天生,周国法.空间自相关与分布型指数研究[J].生态学报,1994,14(3):327-331.
[10]Liebhold A M,Simons E E,Sior A,et al.Geostastical and geographic information system in applied insect ecology[J].Annu.Rev.Entomol,1993,38:303-327.
[11]Rossl R E,Mulla D J,Journel A G,et al.Geostatistical tools formodeling and interpreting ecologicalspatial dependence[J].Ecological Monographs,1992,62(2):277-314.
[12]Futai K,Shirakawa S,Nakai I.The suitability of Korean pine(Pinus koraiensisSieb.et Zucc.)and Japanese Red Pine(P.densiflora Sieb.et Zucc.)as a host of the Japanese pine sawyer,MonochamusalternatusHope(Coleopter: Cerambycid-ae)[J].Appl Ent Zool,1994,29(2):167-177.
[13]IwaoS.A new regression method foranalyzing the aggregation pattern in animal porulations[J].Res Popul Ecol,1968,10:1-20.
[14]TaniwakiT,Okitsu M,KishiY.Diurnalemergence ofMonochamus alternatusHope (Coleoptera:Cerambycidae) from pine logs[J].Journal of the Japanese Forestry Society,2004,89(2):158-163.
[15]雷琼,李孟楼,杨忠歧.花绒坚甲的生物学特性研究[J].西北农林科技大学学报,2003,31(2):62-66.
[16]Miura K,Abe T,Nakashima,Y,et al.Field release of parasitoidDastarcus helophoroides(Fairmaire)(Coleoptera: Bothrideridae)on pine logsinfested withMonochamusalternatusHope (Coleoptera:Cerambycidae)and their dispersal[J].Journal of the Japanese Forestry Society, 2003,85(1):12-17.
[17]Ogura N,Tabata K,Wang W.Rearing of the colydiid beetle predator,Dastarcus helophoroides,on artificial diet [J].BioControl,1999,44(3):291-299.
[18]Urano T.Preliminary release experiments in laboratory and outdoor cages ofDastarcus helophorides(Fairmaire) (Coleoptera: Bothrideridae) for biological control ofMonochamus alternatusHope (Coleoptera:Cerambycidae) [J].Bulletin of the Forestry and Forest Products ResearchInstitute,Ibaraki,2003,389:255-262.
[19]候景儒,黄竞先.地质统计学的理论和方法[M].北京:地质出版社,1990.
[20]Lecoustre L,Fargette D.Analysis and mapping of the spatialspread ofAfrican cassava mosaic virususing geostatisticsand kriging technique [J].Phytopathology, 1989,79:913-920.
Geostastics Analysis of Spatial Patterns of Monochamus Alternatus and Dastarcus Helophoroides
Chen Xiangyang1,2,Zou Yunding2,Bi Shoudong2
(1.School of Life and Environment Science,Huangshan University,Huangshan 245041,China;
2.College of Forestry and Gardening,Anhui Agricultural University,Hefei 230036,China)
Geostastics analysis was used to study the spatial pattern and relationship of monochamus alternatus and its natural enemy-dastarcus helophoroides.The results indicated that the semivariogram of monochamus alternatus and dastarcus helophoroides could be described by spherical model and its spatial construction showed aggregated distribution.The ranges of spatial dependence (RSD)of monochamus alternatus on p.thunbergii lamb and pinus massoniana and p.elliotii in turn were 116.06,33.85,115.76 in 2005;the RSD of monochamus alternatus on p.thunbergii lamb and pinus massoniana in turn were 36.48 and 38.02 in 2004.The ranges of spatial dependence (RSD)of dastarcus helophoroides on p.thunbergii lamb and pinus massoniana and p.elliotii in turn were 92.36,46.50,63.37 in 2005;the RSD of dastarcus helophoroides on p.thunbergii lamb and pinus massoniana in turn were 80.00 and 40.23 in 2004.It could be concluded that their RSDs on the whole were identical and their relationship was the latter on the spacewasafterthe former.Therefore,dastarcushelophoroideswasthe dominantnaturalenemy of monochamus alternatus.
monochamus alternatus;dastarcus helophoroides;spatial pattern;geostastics
Q968.2
A
1672-447X(2010)04-0059-06
2010-02-06
国家自然科学基金重点项目(30330500)
陈向阳(1970-),安徽祁门人,黄山学院生命与环境科学学院讲师,硕士,研究方向为昆虫生态学。
胡德明
