柴达木盆地察尔汗湖区古湖相沉积物中含氮类分子化石的检出
2010-12-14张虎才雷国良常凤琴杨明生
蒲 阳,张虎才,雷国良,常凤琴,杨明生
1)中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,江苏南京 210008;
2)中国科学院研究生院,北京 100049;
3)南昌大学环境与化学工程学院鄱阳湖环境与资源利用教育部重点实验室,江西南昌 330031
柴达木盆地察尔汗湖区古湖相沉积物中含氮类分子化石的检出
蒲 阳1,2),张虎才1),雷国良1),常凤琴1),杨明生3)
1)中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,江苏南京 210008;
2)中国科学院研究生院,北京 100049;
3)南昌大学环境与化学工程学院鄱阳湖环境与资源利用教育部重点实验室,江西南昌 330031
利用气相色谱质谱联用仪(GC-MS)对青藏高原柴达木盆地察尔汗湖区贝壳堤剖面古湖相沉积物样品进行了系统地分析,检测出系列脂肪酸酰胺与脂肪腈类含氮类脂物分子化石。并对其在古湖相沉积物中的分布特征作了详细描述。对它们综合研究认为,早期成岩作用使来源于各种生物体的脂肪酸酰胺类分子化石部分地转化为脂肪腈类化合物。此外,不饱和脂肪酸酰胺与饱和脂肪酸酰胺的含量比值在剖面上并没有呈现明显下降的趋势,说明在降解过程中各种生物、物理及化学作用对它们不存在明显的选择性,指示有机质在沉积过程中可能处于一个偏氧化的环境。本文首次报道这两类物质,为更好地理解含氮类脂物分子化石在湖泊沉积物中的转化过程提供了一定的理论依据。
含氮类分子化石;古湖相沉积物;脂肪酸酰胺;脂肪腈;青藏高原
青藏高原由于其独特的地理位置,已成为第四纪气候、环境变化研究的重点区域(郑绵平等,2008)。同时它也是环境最为脆弱的区域之一,生态系统对气候环境变化的响应极为敏感(陈江等,2007)。而分子化石的优势正是反映生态系统对气候环境变化的响应(Xie et al.,2005)。另一方面青藏高原地区晚第四纪湖泊广泛发育(朱大岗等,2001;赵希涛等,2005),形成了大量的湖泊沉积物,忠实地记录了高原气候环境的变化。因此在不久的将来分子化石手段将会在青藏高原湖泊沉积物研究中广泛应用。
分子化石研究主要涉及蛋白质(包括核酸)、碳水化合物、类脂物和木质素(谢树成等,2003)。其中类脂物分子化石是研究最为广泛的一类分子化石。它们往往具有一定的生物源指示意义,可以直接将它们与特定的生物类别联系起来,从而建立起生物与气候、环境因子的联系(Eglinton and Eglinton,2008)。但是并不是所有类脂物均具有生物的专一性,同时某些来源于生物体的类脂物可能会在成岩,成土过程发生了重大的变化,从而导致类脂物分子化石与气候环境变化的关系比较模糊。因此对各类类脂物分子化石在沉积物中的演化特性及规律进行深入地研究变得十分重要。
非烃类分子化石是指沉积物中来源于生物体的含氧、硫、氮杂原子的有机化合物,每一个饱和烃生物标志化合物都可能对应多个骨架相似,但是包含不同的氧、硫、氮杂原子的非烃类化合物。它们中的许多被认为是沉积物中有机质在向烃类化合物演化过程中的中间产物,其数量庞大,一般极性较大、分离困难。常见的含氧类非烃分子化石大都已被识别,如第四纪沉积物中经常报道的正构脂肪酸/醇系列(Ishiwatari et al.,2006;Dunn et al.,2008;Zhang et al.,2010)、长链烯酮系列(Chu et al.,2005)、五环三萜酮(段毅,2001)、长链酯系列(Morris,1972)等。但是含氮非烃类分子化石研究的相对较少,而在青藏高原湖泊沉积物中还未见报道。生物体中常见的和氮有关的物质主要包括蛋白质及其不同程度的降解产物(肽、氨基酸等)、铵盐、硝酸盐、嘌呤、嘧啶、脲、胺、酰胺和氰化物等。其中脂肪酸酰胺作为一种重要的生物调节剂(bioregulator)形成于生物的诸多生理活动过程中,包括一些生物合成及代谢活动(Bezuglov et al.,1998)。我们对这些来源于生物体的含氮化合物在第四纪沉积物中的分布特征及其转化过程还不是很明确。
以往有关脂肪酸酰胺与脂肪腈的报道主要来自各种生物体(Wu et al.,1997;Kawasaki et al.,1998;Dembitsky et al.,2000;Marketon et al.,2002;Ucar and Ozkan,2008)、气溶胶(Rogge et al.,1991;Simoneit et al.,2003)以及油页岩(Regtop et al.,1982)的研究,而在第四纪沉积物中有关脂肪酸酰胺与脂肪腈类物质的报道非常少.笔者认为这可能主要由于两个原因:①与不同实验室对脂肪酸酰胺类分子化石的检测与鉴定过程有关。Cn脂肪酸酰胺的保留时间与Cn+2脂肪酸接近,利用GC-MS检测时可能会发生共溢出现象,而已有的报告均显示脂肪酸酰胺的相对含量远低于脂肪酸,所以导致Cn脂肪酸酰胺色谱峰在总离子流图上可能被对应的Cn+2脂肪酸“淹没”,从而导致脂肪酸酰胺在很多第四纪沉积物中未被检出。②脂肪酸酰胺和脂肪腈类化合物并不是很稳定,特别是遇水容易水解。所以在现代湖泊中还未有这两类分子化石的报道,同时在中国南方气候相对湿热的环境下也难检测出此类物质。而在青藏高原古湖相沉积物中,这两种含氮类的分子化石在特殊的干旱环境中被相对较好地保存了下来。本文对这两类特殊的分子化石在古湖相沉积物中的分布特征和可能的来源及转化过程进行了详细地报道。
1 研究区域与方法
本次研究的所有样品采自察尔汗古湖贝壳堤剖面,该剖面位于察尔汗古湖东南缘,地理坐标为36°30′N,96°12′E (如图1所示)。本课题组对青藏高原地区柴达木盆地察尔汗湖区贝壳堤剖面已进行了详细的研究(张虎才等,2007a;张虎才等,2007b;Zhang et al.,2008a;Zhang et al.,2008b;张虎才等,2008),对该区域的气候环境变化有了初步的认识。我们认为在43.5至22.4 cal.ka BP之间(相当于深海氧同位素第3阶段晚期)察尔汗古湖经历了湖泊发育早期水位较低→湖面扩张,湖水水位较高→湖面萎缩,水位下降直至湖泊退出贝壳堤所在位置的过程。值得注意的是贝壳堤剖面总有机碳含量在0.05%~0.22%之间变化,平均值为0.11%(常凤琴等,2008),明显低于青藏高原地区现代湖泊的总有机碳含量,如兹格塘错(吴艳宏等,2007)、库赛湖(Liu et al.,2009)等等,甚至低于一些古湖相沉积物中的总有机碳含量(Zhang et al,2004;张成君等,2004)。说明该湖泊在演化过程中生物活动相对微弱,生物种类较少,有机质的来源相对单一。同时也可能说明了在早期成岩过程中,部分有机质被降解。
有关贝壳堤详细的空间展布及剖面描述见文献(张虎才等,2007b)。贝壳堤地层由湖相灰绿色含CaCO3粉砂及粉砂质粘土组成,厚度大且地层特征变化清晰、明显。在野外根据地层变化情况,在260cm的剖面上共采集样品90个。采集到的样品及时冷冻干燥并密封保存。在实验室无尘条件下选取20个样品进行生物标志化合物分析,称取样品 30 g,研磨至100目,采用溶剂-超声波萃取法,将样品用二氯甲烷:甲醇(93:7)混合溶剂浸泡 24 h,再用超声波常温萃取两次,每次 20min,合并萃取液并过滤,将滤液浓缩恒重。由于所得可抽提有机质仅有 5-15mg,为防止样品中微量组分在分离过程中进一步流失,不再用硅胶-氧化铝色谱柱分离为族组分。自然风干并用氯仿稀释后,直接全组分进行 GC-MS分析。所有试剂均为分析纯,并经重新蒸馏后使用。空白样品的GC-MS谱图中未发现文中所要讨论的有机物组分。
有机质分析在中国科学院地质与地球物理研究所气体地球化学重点实验室完成。气相色谱-质谱联用仪型号为HP6890/5973N GC-MS。色谱分析条件:DB-5毛细管柱(30 m×0.25mm×0.25μm),柱始温 80℃,以3℃/min程序升温至300℃,终温恒定20min,进样口温度300℃,载气为氦气。质谱分析条件:电离方式 EI,电离能量 70 eV,离子源温度 230℃,GC-MS接口温度280℃。

图1 研究区域位置示意图(据Chen and Bowler,1986修改)Fig.1 Location of the study area (modified after Chen and Bowler,1986)
2 酰胺类及腈类化合物的检出
图2a为贝壳堤剖面典型样品总离子色谱图。其中所有检测到的脂肪酸酰胺与相对含量较高的脂肪酸分别标注在色谱图上,由于脂肪腈类化合物相对含量较低,在总离子流图上峰形并不明显。表1列出了所有检测出的脂肪酸酰胺与脂肪腈系列的分子式及其它相关信息。图2b是总离子流提取碎片离子72后的质量色谱图,系列脂肪酸酰胺标注在图上。可以发现脂肪酸酰胺具有明显的偶碳优势,碳数分布范围为C14~C20,奇碳数的脂肪酸酰胺在样品中几乎检测不到。图2c是总离子流提取碎片离子97后的质量色谱图,脂肪腈可以看作氢氰酸HCN的氢原子被脂肪烃基取代而生成的化合物,通式为R-C≡N。对比图 2b、c可以发现系列脂肪腈的分布与系列脂肪酸酰胺具有相似的碳数分布模式,表现在均呈现相同的碳数分布范围,主峰碳数为18且含有对应的不饱和脂肪酸酰胺和脂肪腈。
脂肪酸酰胺的谱图以59,72为基峰,同时结合分子离子峰及同系物等间距出峰的特征,以及与标准图谱较高的匹配率,分别确定出 38.38、44.43、50.94分钟所出色谱峰分别为正十四酰胺,正十六酰胺以及正十八酰胺。由于单不饱和脂肪酸酰胺比饱和脂肪酸酰胺多一个双键,导致其离子碎片相应减2Da(图3),据此可确定 9-十八烯酰胺。同理通过与标准谱库中质谱图及文献27的比对(Simoneit et al.,2003),确定30.16、35.92、42.16分钟所出的色谱峰分别为正十四腈,正十六腈及正十八腈。NIST谱库中没有给出十八烯腈的标准图谱,所以根据保留时间以及离子碎片分布特征(图3),判断在 41.40分钟时所出色谱峰为9-十八烯腈。

图2 典型样品总离子流图(a)、提取特征离子72后的质量色谱图(b)、提取特征离子97后的质量色谱图(c)、提取特征离子73后的质量色谱图(d)(图b和c中所标注数字与表1峰号对应)Fig.2 GC-MS mass chromatogram of total ion current,fatty acid amide (m/z 72),fatty nitrile (m/z 97)and n-fatty acids (m/z 73)identified in lake sediments

图3 9-十八烯酰胺(a)、正十八酰胺(b)、9-十八烯腈(b)、正十八烯腈质谱图(d)Fig.3 Mass spectrometry of 9-Octadecenamide(a),Octadecanamide(b),9-Octadecenenitrile(c)and Octadecanenitrile(d)

表1 察尔汗古湖贝壳堤剖面古湖相沉积物样品中检测出的脂肪酸酰胺与脂肪腈名称及基本参数表Table 1 Related information of n-fatty acid amide and fatty n-nitrile in lake sediments of the shell bar section,Qaidam Basin
3 酰胺类及腈类化合物在贝壳堤剖面中可能的来源及转化过程
3.1 脂肪酸酰胺可能的来源及转化
对比脂肪酸酰胺与脂肪酸的碳数分布特征,可以发现它们之间存在明显的差异。脂肪酸碳数分布在C12至C18之间且存在明显的奇碳数脂肪酸,以正十六脂肪酸为主峰(图 2d);而脂肪酸酰胺碳数分布在C14至C20之间,仅检测到偶碳数系列的脂肪酸酰胺,以正十八脂肪酸酰胺为主峰(图2b)。这两类化合物碳数分布明显的差异性。Simoneit et al.(2003)为了研究气溶胶中烷基酸胺的来源,将脂肪酸和(NH4)HCO3混和装入高压釜,加热至300℃反应72小时后得到脂肪酸酰胺与脂肪腈。由于其具有明显的偶碳优势及相似的碳数分布特征,推测脂肪酸酰胺是由脂肪酸和氨或氨离子(可能来自蛋白质降解)反应得到,然后在高温条件下进一步生成脂肪腈。由于在本次研究同一样品中的脂肪酸与脂肪酸酰胺的碳数分布特征迥异,说明脂肪酸与脂肪酸酰胺的来源可能不同,意味着察尔汗古湖相沉积物中脂肪酸酰胺并不是从脂肪酸经过酰胺化过程转化过来。
从脂肪酸酰胺的碳数分布形态可判断它们主要来源于生物体,而不是各种可能的污染源(如化学试剂或者石油残余物),主要的原因是:①脂肪酸酰胺系列具有强烈的偶碳优势;②存在相对丰度较高的不饱和脂肪酸酰胺(C18:1)。③未检测出碳数高于20的高碳数脂肪酸酰胺。已有的报道显示各种生物在高温或者燃烧的条件下,可以产生高碳数脂肪酸酰胺,这类脂肪酸酰胺的分布特征是以高碳数不饱和脂肪酸酰胺(如 C20:1、C22:1)为主峰(Simoneit et al.,2003)。
前人的研究成果显示在各种生物体内均可能含有系列脂肪酸酰胺类化合物。比如神经酰胺、鞘脂类、酰化类脂物、脂蛋白等物质中均含有脂肪酸酰胺的结构(Bezuglov et al.,1998)。在生物体中它们往往是在脂肪酸与胺类化合物在酰胺化作用下形成的。首先是在很多高等植物中检测到了长链脂肪酸酰胺:在油菜中检测到了正十六与正十八酰胺及 9-十八烯酰胺(Ucar and Ozkan,2008)。海洋草本植物大叶藻(Zostera marina)中可挥发有机组分中检测到了正十六酰胺(1.1%)与正十八酰胺(1.2%)(Kawasaki et al.,1998)。各种微生物和藻类中也检测到多种脂肪酸类酰胺:淡水绿藻根枝藻(Rhizoclonium hieroglyphicum)中检测到八种脂肪酸酰胺,包括从 C4到C18的饱和与不饱和脂肪酸酰胺(Dembitsky et al.,2000);生活在海洋中的巨大鞘丝藻(Lyngbya majuscula)中检测到三种 malyngamide类酰胺化合物,被认为是海洋藻类新陈代谢的产物(Wu et al.,1997);苜蓿根瘤菌中检测到了被认为是来源于细胞膜的C12-C18的N-酰基-高丝氨酸内酯(AHLs),而此类物质非常容易水解生成对应的酰胺物质(Marketon et al.,2002);海洋原生动物有毒甲藻(Coolia monotis)中检测到支链脂肪酸酰胺(Tanaka et al.,2000)。虽然脂肪酸酰胺存在于不同类型的生物体中,但是其种类及分布模式存在很大的差异,所以此类化合物具备成为灵敏指示生物种类的分子化石的条件。
基于前人和本研究的分析结果,我们认为柴达木盆地贝壳堤剖面中检测到系列长链正脂肪酸酰胺及不饱和长链脂肪酸酰胺直接来源于湖泊周围及湖泊内部的各种生物体,包括高等植物、水生藻类以及各种微生物,反映的是一个混合信息,并不能确定其具体的生物来源。同时某些来源于生物体的类脂物可能会在成岩/成土过程发生了明显的变化,导致类脂物分子化石与古气候、古环境变化的关系比较模糊。所以我们认为难以精确提取其中的古环境、古气候信息。但是有学者认为除了直接来源于生物体且具有一定生物专一性的类脂物分子具有古气候与古环境意义外,那些受后期改造的地质类脂物也可以记录过去全球变化的信息(谢树成等,2008)。所以脂肪酸酰胺类分子化石的古气候、古环境意义有待于进一步的研究。
有意义的是柴达木盆地贝壳堤剖面不饱和脂肪酸酰胺相对饱和脂肪酸酰胺的量并没有出现明显下降的趋势,甚至底层样品中不饱和脂肪酸酰胺的相对量比表层样品的还要高(见图4)。这一现象可与一些湖泊沉积物剖面中脂肪酸的变化趋势进行类比。如日本琵琶湖剖面不饱和脂肪酸和饱和脂肪酸的比值在剖面上相对稳定,并未出现随剖面深度的加深不饱和脂肪酸相对含量降低的特征(Kawamura et al.,1980)。此类湖泊的特征是水体相对较大,水位较深,有机质在沉降的过程中处于一个偏氧化的环境中,而沉积到湖泊沉积物中之后保存相对较好,可记录古气候信息(Kawamura and Ishiwatari,1981)。一般认为在还原条件下,不饱和脂肪酸由于不饱和双键的存在导致其更容易受到微生物的降解;而在氧化环境中不饱和脂肪酸与饱和脂肪酸的降解速率相当。同样的机理在不饱和脂肪酸酰胺与饱和脂肪酸酰胺之间也应存在。从贝壳堤样品中存在高相对含量的不饱和脂肪酸酰胺可以推断察尔汗古湖的演化过程中曾经经历过湖泊面积较大,水位较深的阶段,沉积环境为偏氧化环境。脂肪酸酰胺在剖面上的分布特征所反映出来的湖泊演化特征与其它地质记录并不矛盾(张虎才等,2007a;张虎才等,2007b;Zhang et al.,2008a;Zhang et al.,2008b;张虎才等,2008)。
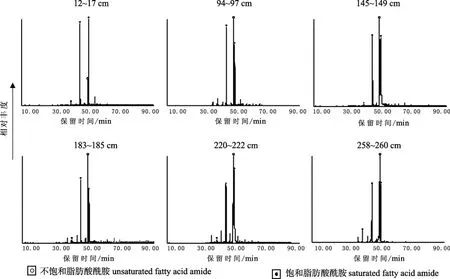
图4 贝壳堤剖面不同层位脂肪酸酰胺碳数分布(m/z 72)Fig.4 Distribution of n-fatty acid amide in different layers of the shell bar section,Qaidam Basin (m/z 72)
3.2 脂肪腈在古湖相沉积物中的来源
短链的腈类化合物(碳链长度小于等于 6)可直接来源于高等植物体,如在油菜(Radovic et al.,2001)、各种单花种蜂蜜(Fuente et al.,2007)中均可以检测到系列短链脂肪腈。同时短链的脂肪腈类化合物容易被微生物改造,生成对应的酰胺和羧酸。另一方面在天然产物中未见有长链脂肪腈的报道。而在页岩油(shale oil)中大量出现长链脂肪腈类化合物(质量百分比最高可达 13%),并且与同一样品中的脂肪酸同系物具有相似的分布模式,从而推断这些脂肪酸腈是在高温环境下由脂肪酸与胺类物质反应后生成(Evans et al.,1985)。从总离子流图(图2a)上可以发现系列脂肪酸酰胺的相对含量明显高于系列脂肪腈。而脂肪腈与脂肪酸酰胺的分布模式比较接近,且所有脂肪酸酰胺均有对应的脂肪腈类化合物。所以我们认为柴达木盆地贝壳堤剖面具有明显奇偶优势的长链饱和及不饱和腈类化合物不可能直接来源于生物体,也不可能来源于石油残余物的污染,而是来源于生物体的酰胺类化合物经过早期的成岩作用而逐渐形成脂肪腈类物质。分子有机地球化学上的成岩作用(diagenesis)与沉积学中的成岩作用明显不同。有机分子从离开生物体后进入地质体并最终保存下来的演变过程,是按成岩作用、深成作用(catagenesis)和变成作用(metagenesis)过程来划分的。生物有机分子从离开生物体,到深成作用之前所发生的生物、物理以及化学变化的过程都属于成岩作用(谢树成等,2007)。古湖相沉积物中长链脂肪腈的检出说明早期成岩作用对湖泊沉积物中的脂肪酸酰胺类分子化石已产生了显著的影响,在进行相关研究时需加以考虑。
4 结论
柴达木盆地贝壳堤剖面含有系列脂肪酸酰胺与脂肪腈化合物,这两个系列的化合物均呈现了相似的碳数分布模式,并且都含有对应的不饱和脂肪酸酰胺和脂肪腈。脂肪酸酰胺强烈的偶碳优势及高相对含量的不饱和脂肪酸酰胺的检出说明此类化合物主要来源于生物体,并被较好地保存于古湖相沉积物中。不饱和脂肪酸酰胺与脂肪酸酰胺相对量的比值并没有在剖面上呈现逐渐下降的趋势,说明微生物对不饱和脂肪酸酰胺和饱和脂肪酸酰胺的改造程度相似,而不存在选择性降解。由于脂肪腈类化合物与脂肪酸酰胺类化合物具有相似的碳数分布特征,同时结合前人的研究结论,我们认为贝壳堤剖面古湖相沉积物中脂肪腈类化合物可能主要是由脂肪酸酰胺经早期成岩作用改造后所生成。
常凤琴,张虎才,陈玥,杨明生,牛洁,樊红芳,雷国良,张文翔,类延斌,杨伦庆.2008.柴达木盆地察尔汗古湖贝壳堤剖面沉积地球化学与环境变化[J].地球科学,33(2):197-204.
陈江,万力,梁四海,金晓媚,陈立 2007.青藏高原生态环境变化趋势的初步探索[J].地球学报,28(6):555-560.
段毅.2001.甘南沼泽泥炭中五环三萜酮系列化合物的检出[J].科学通报,46(11):960-962.
吴艳宏,LÜCKE,A,WÜNNEMANN,B,李世杰,王苏民.2007.青藏高原中部全新世气候变化的湖泊沉积地球化学记录[J].中国科学(D辑):地球科学,37(9):1185-1191
谢树成,赖旭龙,黄咸雨,马相如,杨淑娟.2007.分子地层学的原理、方法及应用实例[J].地层学杂志,31(3):209-221.
谢树成,梁斌,顾延生,杨欢.2008.脂肪酮分子在第四纪古土壤中的分布及其古气候意义[J].古生物学报,47(3):273-278.
谢树成,梁斌,郭建秋,易轶,EVERSHED R P,MADDY,D,CHAMBERS F M.2003.生物标志化合物与相关的全球变化[J].第四纪研究,23(5):521-528.
张成君,陈发虎,尚华明,曹洁.2004.中国西北干旱区湖泊沉积物中有机质碳同位素组成的环境意义—以民勤盆地三角城古湖泊为例[J].第四纪研究,24(1):88-94.
张虎才,常凤琴,李斌,雷国良,陈玥,张文翔,牛洁,樊红芳,杨明生.2007a.柴达木察尔汗湖贝壳堤剖面长链支链烷烃及其古环境意义[J].科学通报,52(6):707-714.
张虎才,雷国良,常凤琴,樊红芳,杨明生,张文翔.2007b.柴达木盆地察尔汗贝壳堤剖面年代学研究[J].第四纪研究,27(4):511-521.
张虎才,王强,彭金兰,陈光杰.2008.柴达木察尔汗盐湖贝壳堤剖面介形类组合及其环境意义[J].第四纪研究,28(1):103-111.
赵希涛,吴中海,胡道功,严富华,马志邦,麦学舜,鄢犀利.2005.西藏错鄂及邻区晚更新世高位湖相沉积的发现及其意义[J].地球学报,26(4):291-298.
郑绵平,刘喜方,袁鹤然,张成君,王海雷.2008.青藏高原第四纪重点湖泊地层序列和湖相沉积若干特点[J].地球学报,29(3):293-305.
朱大岗,赵希涛.2001.纳木错湖相沉积与藏北高原古大湖[J].地球学报,22(2):149-155.
BEZUGLOV V,BOBROV M,ARCHAKOV A.1998.Bioactive amides of fatty acids[J].Biochemistry (Moscow),63(1):22-30.
CHANG Feng-qin,ZHANG Hu-cai,CHEN Yue,YANG Ming-sheng,NIU Jie,FAN Hong-fang,LEI Guo-liang,ZHANG Wen-xiang,LEI Yan-bing,YANG Lun-qing.2008.Sedimentation Geochemistry and Environmental Changes during the Late Pleistocene of Paleolake Qarhan in the Qaidam Basin[J].Journal of China University of Geosciences,33(2):197-204(in Chinese).
CHEN Jiang,WAN Li,LIANG Si-hai,JIN Xiao-mei,CHEN Li.2007.A tentative discussion on the trend of ecological environment change in Qinghai-Tibet Plateau[J].Acta Geoscientica Sinica,28(6):555-560(in Chinese).
CHEN K Z,BOWLER J M.1986.Late Pleistocene evolution of salt lakes in the Qaidam Basin,Qinghai Province,China[J].Palaeogeography,Palaeoclimatology,Palaeoecology,54:87-104.
CHU Guo-Qiang,SUN Qing,LI Sheng-Qiang,ZHENG Mian-Ping,JIA Xin-Xian,LU Cai-fen,LIU Jia-qi,LIU Tung-sheng.2005.Long-chain alkenone distributions and temperature dependence in lacustrine surface sediments from China[J].Geochimica et Cosmochimica Acta,69(21):4985-5003.
FUENTE D L,SANZ M L,MARTÍEZ-CASTRO I,SANZ J,RUIZ-MATUTE A I.2007.Volatile and carbohydrate composition of rare unifloral honeys from Spain[J].Food Chemistry,105(1):84-93.
DEMBITSKY V M,SHKROB I,ROZENTSVET O A.2000.Fatty acid amides from freshwater green alga Rhizoclonium hieroglyphicum[J].Phytochemistry,54(8):965-967.
DUAN Yi.2001.Pentacyclic triterpenoid ketones in peat from Gannan Marsh[J].Chinese Science Bulletin,46(17):1433-1436.
DUNN R J K,WELSH D T,TEASDALE P R,LEE S Y,LEMCKERT C J,MEZIANE T.2008.Investigating the distribution and sources of organic matter in surface sediment of Coombabah Lake (Australia)using elemental,isotopic and fatty acid biomarkers[J].Continental Shelf Research,28(18):2535-2549.
EGLINTON T I,EGLINTON G.2008.Molecular proxies for paleoclimatology[J].Earth and Planetary Science Letters,275(1-2):1-16.
EVANS E J,BATTS B D,CANT N W,SMITH J W.1985.The origin of nitriles in shale oil[J].Organic Geochemistry,8(5):367-374.
ISHIWATARI R,YAMAMOTO S,SHINOYAMA S.2006.Lignin and fatty acid records in Lake Baikal sediments over the last 130Kyr:A comparison with pollen records[J].Organic Geochemistry,37(12):1787-1802.
KAWAMURA K,ISHIWATARI R,YAMAZAKI M.1980.Identification of polyunsaturated fatty acids in surface lacustrine sediments[J].Chemical Geology,28:31-39.
KAWAMURA K,ISHIWATARI R.1981.Polyunsaturated fatty acids in a lacustrine sediment as a possible indicator of paleoclimate[J].Geochimica et Cosmochimica Acta,45(2):149-155.
KAWASAKI W,MATSUI K,AKAKABE Y,ITAI N,KAJIWARA T.1998.Volatiles from Zostera marina[J].Phytochemistry,47(1):27-29.
LIU X Q,DONG H L,YANG X D,HERZSCHUH U,ZHANG E L,STUUT J-B W,WANG Y B.2009.Late Holocene forcing of the Asian winter and summer monsoon as evidenced by proxy records from the northern Qinghai-Tibetan Plateau[J].Earth and Planetary Science Letters,280(1-4):276-284.
MARKETON M,GRONQUIST M,EBERHARD A,GONZALEZ J.2002.Characterization of the Sinorhizobium meliloti sinR/sinI locus and the production of novel N-acyl homoserine lactones[J].Journal of Bacteriology,184(20):5686-5695.
MORRIS R J.1972.The occurrence of wax esters in crustaceans from the North-east Atlantic Ocean[J].Marine Biology,16(2):102-107.
RADOVIC B S,CARERI M,MANGIA A,MUSCI M,GERBOLES M,ANKLAM E.2001.Contribution of dynamic headspace GC-MS analysis of aroma compounds to authenticity testing of honey[J].Food Chemistry,72(4):511-520.
REGTOP R A,CRISP P T,ELLIS J.1982.Chemical characterization of shale oil from Rundle,Queensland[J].Fuel,61(2):185-192.
ROGGE W F,HILDEMANN L M,MARUREK,M A,CASS,G R.1991.Sources of fine organic aerosol.1.Charbroilers and meat cooking operations[J].Environmental Science Technology,25(6):1112-1125.
SIMONEIT B R T,RUSHDI A I,BINABAS M R,DIDYK B M.2003.Alkyl Amides and Nitriles as Novel Tracers for Biomass Burning[J].Environment Science Technology,37(1):16-21.
TANAKA I,MATSUOKA S,MURATA M,TACHIBANA K.2000.Absolute configuration of a ceramide with a novel branched-chain fatty acid isolated from the epiphytic dinoflagellate,Coolia monotis[J].Bioscience,Biotechnology,and Biochemistry,64(9):1842-1846.
UCAR S,OZKAN A R.2008.Characterization of products from the pyrolysis of rapeseed oil cake[J].Bioresource Technology,99(18):8771-8776.
WU M,MILLIGAN K E,GERWICK W H.1997.Three new malyngamides from the marine cyanobacterium Lyngbya majuscula[J].Tetrahedron,53(47):15983-15990.
WU Yan-hong,LÜCKE ANDREAS,WÜNNEMANN BERND,LI Shi-Jie,WANG Su-min.2007.Holocene climate change in the Central Tibetan Plateau inferred by lacustrine sediment geochemical records[J].Science in China series D:Earth Science,50 (10):1548-1555.
XIE S C,PANCOST R D,YIN H F,WANG H M,EVERSHED R P.2005.Two episodes of microbial change coupled with Permo/Triassic faunal mass extinction[J].Nature,434(24):494-497.
XIE Shu-cheng,LAI Xu-long,HUANG Xian-yu,MA Xiang-ru,YANG Shu-juan.2007.Principles,methodology and application of molecular stratigraph[J].Journal of Stratigraphy,31(3):209-221(in Chinese).
XIE Shu-cheng,LIANG Bing,GU Yan-sheng,YANG Huan.2008.Distributions of n-alkan-2-ones in quaternary paleosols indicative of paleoclimate changes[J].Acta Palaeontologica Sinica,47(3):273-278(in Chinese).
XIE Shu-cheng,LIANG Bing,GUO Jian-qiu,YI Yi,EVERSHED R P,MADDY D,CHAMBERS F M.2003.Biomarkers and the related global change[J].Quaternary Science,23(5):521-528(in Chinese).
ZHANG Cheng-jun,CHEN Fa-hu,SHANG Hua-ming,CAO Jie.2004.The paleoenvironmental significance of organic carbon isotope in lacustrine sediments in the arid China:an experimental from Sanjiaocheng palaeolake in Minqin[J].Quaternary Science,24(1):88-94(in Chinese).
ZHANG Hong-liang,LI Shi-jie,FENG Qing-lai,ZHANG Shi-tao.2010.Quaternary International.Environmental change and human activities during the 20th century reconstructed from the sediment of Xingyun Lake,Yunnan Province,China[J].212(1):14-20.
ZHANG Hu-Cai,FAN Hong-Fang,CHANG Feng-Qin,ZHANG Wen-Xiang,LEI Guo-Linag,YANG Ming-Sheng,LEI Yan-Bin,YANG Lun-Qing.2008a.AMS dating on the Shell Bar section from Qaidam Basin,NE Tibetan Plateau,China[J].Radiocarbon,50(2):255-265.
ZHANG Hu-Cai,LEI Guo-Liang,CHANG Feng-Qin,PU Yang,FAN Hong-Fang,LEI Yan-Bin,YANG Ming-Sheng,ZHANG Wen-Xiang,YANG Lun-Qing,2008b.Chronology of the shell bar section and a discussion on the ages of the Late Pleistocene lacustrine deposits in the paleolake Qarhan,Qaidam basin[J].Frontiers of Earth Science in China,2(2):225-235.
ZHANG Hu-cai,CHANG Feng-qin,LI Bing,LEI Guo-liang,CHEN Yue,ZHANG Wen-xiang,NIU Jie,FAN Hong-fang,YANG Ming-sheng.2007.Branched aliphatic alkanes of the Late Pleistocene paleolake deposits from the Qaidam Basin,NE Tibetan Plateau[J].Chinese Science Bulletin,52(9):1248-1256.
ZHANG Hu-cai,LEI Guo-liang,CHANG Feng-qin,FAN Hong-fang,YANG Ming-sheng.ZHANG Wen-xiang.2007b.Age Determination of shell bar section in salt lake Qarhan,Qaidam Basin[J].Quaternary Science,27(4):511-521(in Chinese).
ZHANG Hu-cai,WANG Qiang,PENG Jin-lan,CHEN Guang-jie.2008.Ostracod assemblages and their paleoenvironmental significance from shell bar section of paleolake Qarhan,Qaidam basin[J].Quaternary Science,28(1):103-111(in Chinese).
ZHANG Z H,ZHAO M X,YANG X D,WANG S M,JIANG X Z,FRANK O,GEOFFREY E.2004.A hydrocarbon biomarker record for the last 40 kyr of plant input to Lake Heqing,southwestern China[J].Organic Geochemistry,35(5):595-613.
ZHAO Xi-tao,WU Zhong-hai,HU Dao-gong,YAN Fu-hua,MA Zhi-bang,MAI Xue-shun,YAN Xi-Li.2005.The discovery of late Pleistocene highstand lacustrine sediments of the Co Ngoin Lake and adjacent areas,Tibet[J].Acta Geoscientica Sinica,26(4):291-298(in Chinese).
ZHENG Mian-ping,LIU Xi-fang,YUAN He-ran,ZHANG Cheng-jun,WANG Hai-lei.2008.Some Characteristics of stratigraphic sequences and lacustrine sediments of main quaternary lakes on the Qinghai-Tibet Plateau[J].Acta Geoscientica Sinica,29(3):293-305(in Chinese).
ZHU Da-gang,ZHAO Xi-tao.2001.Nam Tso lacustrine sediments and the ancient big lake in northern Tibet Plateau[J].Acta Geoscientica Sinica,22(2):149-155(in Chinese).
The Distribution of Nitrogen-bearing Lipid Biomarker in the Shell Bar Paleolake Sediments from Qarhan Paleolake,Qaidam Basin
PU Yang1,2),ZHANG Hu-cai1),LEI Guo-liang1),CHANG Feng-qin1),YANG Ming-sheng3)
1)State Key Laboratory of Lake Science and Environment,Nanjing Institute of Geography and Limnology,Chinese Academy of Sciences,Nanjing,Jiangsu210008;
2)Graduate University of Chinese Academy of Sciences,Beijing100049;
3)Key Laboratory of Boyang Lake,MOE,School of Environmental and Chemical Engineering,Nanchang Universiy,.Nanchang,Jiangxi330031
A series of analogs of fatty acid amide and fatty nitrile were identified along the shell bar section of the Qaidam Basin in Qinghai-Tibet Plateau by using the gas chromatography-mass spectrometry (GC-MS).On such a basis,the distribution characteristics of the nitrogen-bearing lipids have been discussed in detail in this paper.According to previous studies and geographical conditions of the Qaidam Basin,the authors hold that the early diagenesis partly converted the fatty acid amide into the fatty nitrile.However,the ratio of the unsaturated fatty acid amide to the saturated one is relatively stable in the profile,indicating that there existed no obvious selectivity between these two types of fatty acid amides during the diagenetic processes including various biological,chemical and physical actions.It is therefore concluded that the depositional environment in the Qarhan paleolake was relatively aerobic during the organic component deposition.Being the first description of the fatty acid amide and fatty nitrile in paleolake sediments,this report provides the basis for the nitrogen-bearing lipids investigation in lake sediments.
nitrogen-bearing molecular fossil;paleolake sediments;fatty acid amide;fatty nitrile;Qinghai-Tibet plateau
O623.1;O623.76
A
1006-3021(2010)02-155-08
本文由国家自然科学基金项目(编号:40871096)和中国科学院“百人计划”项目联合资助。
2010-01-21;改回日期:2010-04-08。
蒲阳,男,1980年生。博士研究生。主要从事自然地理学和生物标志化合物在全球变化中的应用研究。通讯地址:210008,南京市北京东路73号。电话:025-86882091。E-mail:puyangnew@126.com。
致谢:感谢王有孝研究员、王永莉博士对样品的前期处理及分析工作。
