应用分子标记探讨多胚亚麻的分类地位
2010-12-05康庆华黄文功姜卫东赵东升宋喜霞孙中义吴广文关凤芝
康庆华 ,黄文功 ,刘 岩 ,姜卫东 ,赵东升 ,宋喜霞 ,孙中义 ,吴广文 ,关凤芝
(1.黑龙江省农业科学院经济作物研究所,哈尔滨150086;2.黑龙江省农业科学院畜牧研究中心,哈尔滨 150086)
多胚亚麻品系1-6Ha-3是从俄罗斯引进的多胚种质D95029(形态特征接近油用亚麻)与纤维亚麻抗6的杂交后代中多胚种子产生的单倍体苗加倍后获得的双单倍体(DH)品系。该品系形态特征明显区别于油用亚麻,更接近纤维亚麻,而且株高、工艺长等农艺性状都优于亲本。经两年的所内鉴定,其原茎产量、纤维产量、全麻率都高于对照品种黑亚11号和黑亚14号,同时具有高频率的多胚特性。为直接用于生产或作为优良育种亲本更好的利用,明确其在栽培亚麻品种中的地位及与各品种间的遗传差异非常必要。
目前,国内外已经有很多成功应用RAPD(随机扩增多态性)技术和SSR(简单序列重复,又称微卫星标记)等分子标记技术探讨物种的分类及亲缘关系的研究报道[1-8]。随后,基于微卫星序列发展起来的ISSR(inter-simple sequence repeat,即简单序列重复区间扩增多态性)技术,因其具有重复性好、多态性高等优点,也被广泛用于作物遗传多样性研究、种质资源的分类和鉴定、重要农艺基因的定位与遗传作图上[9-10]。本研究以RAPD技术和ISSR技术相结合的方法对所获得的多胚DH品系1-6Ha-3与其他23个亚麻品种的亲缘关系进行了分析,旨在为多胚亚麻品种的鉴定、保护和分子标记辅助育种提供理论依据。同时也为筛选亚麻多胚性分子标记的研究工作奠定基础。
1 材料与方法
1.1 材料
试验于2008年6月至2009年1月在东北农业大学分子生物实验室进行。供试24个亚麻品种(系)均由黑龙江省农业科学院经济作物研究所提供,见表1。
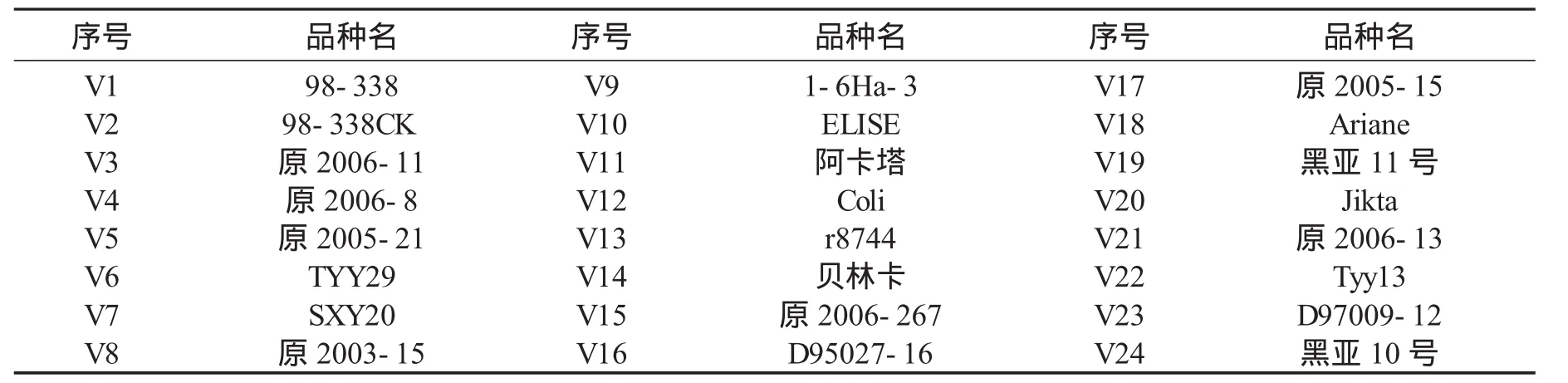
表1 供试的24份亚麻材料Table 1 Twentyfour materials ofLinum usitatissmum L.for the experiment
1.2 方法
1.2.1 总DNA的提取:亚麻总DNA的提取试验参考王关林、方宏筠的方法[11],取0.2g亚麻幼嫩叶片,加500 μL CTAB(内含0.4%巯基乙醇和1%PVP),用液氮研磨,利用购自天根生化科技(北京)有限公司的DNA提取试剂盒提取。
1.2.2 PCR扩增体系及程序:按黄文功(2009)的方法[12]进行。
1.2.2.1 引物:供试引物包括RAPD随机引物70条:A1~A16,S1~S50,S61~S64;ISSR引物21条,选自丁明忠等(2009)在苎麻分子标记中筛选到的U系列引物[13]。以上引物均购自上海生工。
1.2.2.2 PCR 反应体系:总体积 25 μL:10×Buffer 2.5 μL,25 mmol/LMgCl21.0 μL,2.5 mmol/LdNTP 2.5μL,primer(25μmol/L)1.5μL,GenomeDNA2μL(50ng),5U/μLTaq酶0.2μL,加ddH2O至总体积25μL。Marker:100bpladder。以上常规试剂均购自天根生化科技(北京)有限公司。
1.2.2.3 反应程序:94℃预变性4 min;然后94℃变性40 s,37℃退火1 min,72℃延伸90 s,共40个循环;72℃延伸10 min。扩增产物采用0.8%琼脂糖凝胶,80 V电压下电泳。EB(溴化乙锭)染色,紫外分析仪上检测并照相。
1.2.3 数据处理与统计:综合各引物的扩增图谱,筛选在这24份材料中扩增丰富且多态性好的引物。各扩增产物的记录以所用引物加上其相对分子量大小表示,每一条带在某一个样品存在赋值“1”,缺乏赋值“0”,强带和可重复的弱带赋值均为“1”。数据录入NTSYSpc2.10e软件,计算各个样品间的遗传相似性系数(GS),对得到的遗传相似性矩阵进行非加权组法(UPGMA)聚类分析,建立该多胚亚麻品系与其他供试品种(系)间的亲缘关系图。根据遗传相似系数计算出遗传距离GD(遗传距离=1-遗传相似度)[14]。
2 结果与分析
2.1 引物的筛选
选择一定数量的引物对供试材料进行扩增是获得客观结果的前提。本试验在随机选择的70条RAPD引物和21条ISSR引物中筛选出对供试亚麻材料能扩增出清晰的谱带、重复性好、多态性丰富的引物10条,其中RAPD引物5条,ISSR引物5条(见表2)。
2.2 亚麻DNA的PCR扩增结果
2.2.1 24个亚麻品种的DNA多态性
筛选出的10条引物扩增的带型较理想,多态性较好,3次重复扩增的结果一致。这10个引物共扩增出115条带,其中同源片段28条,多态性条带87条,多态性比例为75.7%。平均每条引物扩增出11.5条带,谱带大小一般在200~2000bp,也有极少数在100bp~200bp之间或超过2000bp。其中扩增谱带最多的引物是U835(15条),扩增出的谱带最少的引物有A5和U853,各为9条。表2为10条引物在24个亚麻品种中的扩增结果。图1、图2和图3为选用3个引物A12、A5和U853的基因组DNA的指纹图谱。

表2 10条引物的RAPD和ISSR扩增结果Table 2 Ten primers and their RAPDand ISSR amplification results
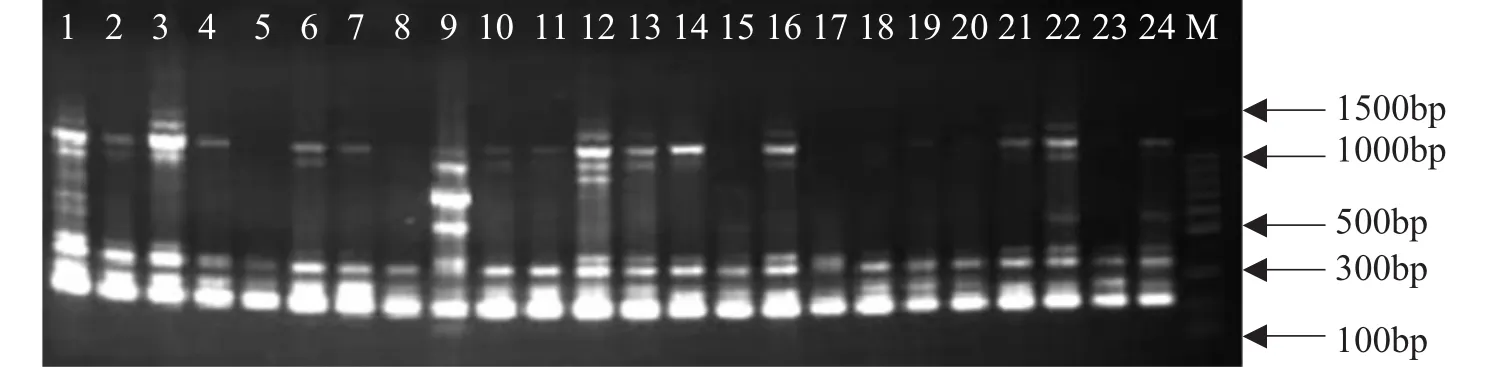
图1 引物A12在24个亚麻品种中的PCR扩增结果1-24.亚麻品种编号(表 1);M.100bp ladderFig.1 PCR amplification results ofthe 24 flaxcultivars usingprimer A121-24:flaxvarieties sample number(table 1);M:100bp ladder
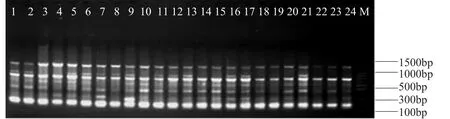
图2 引物A5在24个亚麻品种中的PCR扩增结果1-24.亚麻品种编号(表 1);M.100bp ladderFig.2 PCR amplification results ofthe 24 flaxcultivars usingprimer A51-24:flaxvarieties sample number(table 1);M.100bp ladder

图3 引物U853在24个亚麻品种中的PCR扩增结果1-24.亚麻品种编号(表 1);M.100bp ladderFig.3 PCR amplification results ofthe 24 flaxcultivars usingprimer U8531-24:flaxvarieties sample number(table 1);M.100bp ladder
2.2.2 聚类分析结果
根据10个引物的扩增条带的统计结果进行聚类分析,得到24个亚麻品种(系)的遗传相似度值(表3)和遗传聚类树状图(图4)。24份亚麻种质在阈值(遗传相似系数)为0.73处分为四大类,V19(黑亚 11 号)、V18(Ariane)、V23(D97009-12)等 21 品种聚为第一大类,1-6Ha-3、黑亚 10 号、原05-15分别为第二、三、四类群。1-6Ha-3与其他23个亚麻品种的遗传相似系数范围分布在0.6522~0.7739之间,遗传距离由大到小为V24/V21/V19>V23>V17>V15/V4>V16>V20>V13>V3>V22>V14>V6>V18>V12>V2>V8>V10>V11>V7>V1/V5。遗传距离计算公式得:1-6Ha-3与黑亚 10号(V24)、原 2006-13(V21)、黑亚 11号(V19)三个品种的遗传距离最远,都为 0.3478,与品种原 2005-21 白(V5)、98-338(V1)的遗传距离相对最近,为 0.2261。
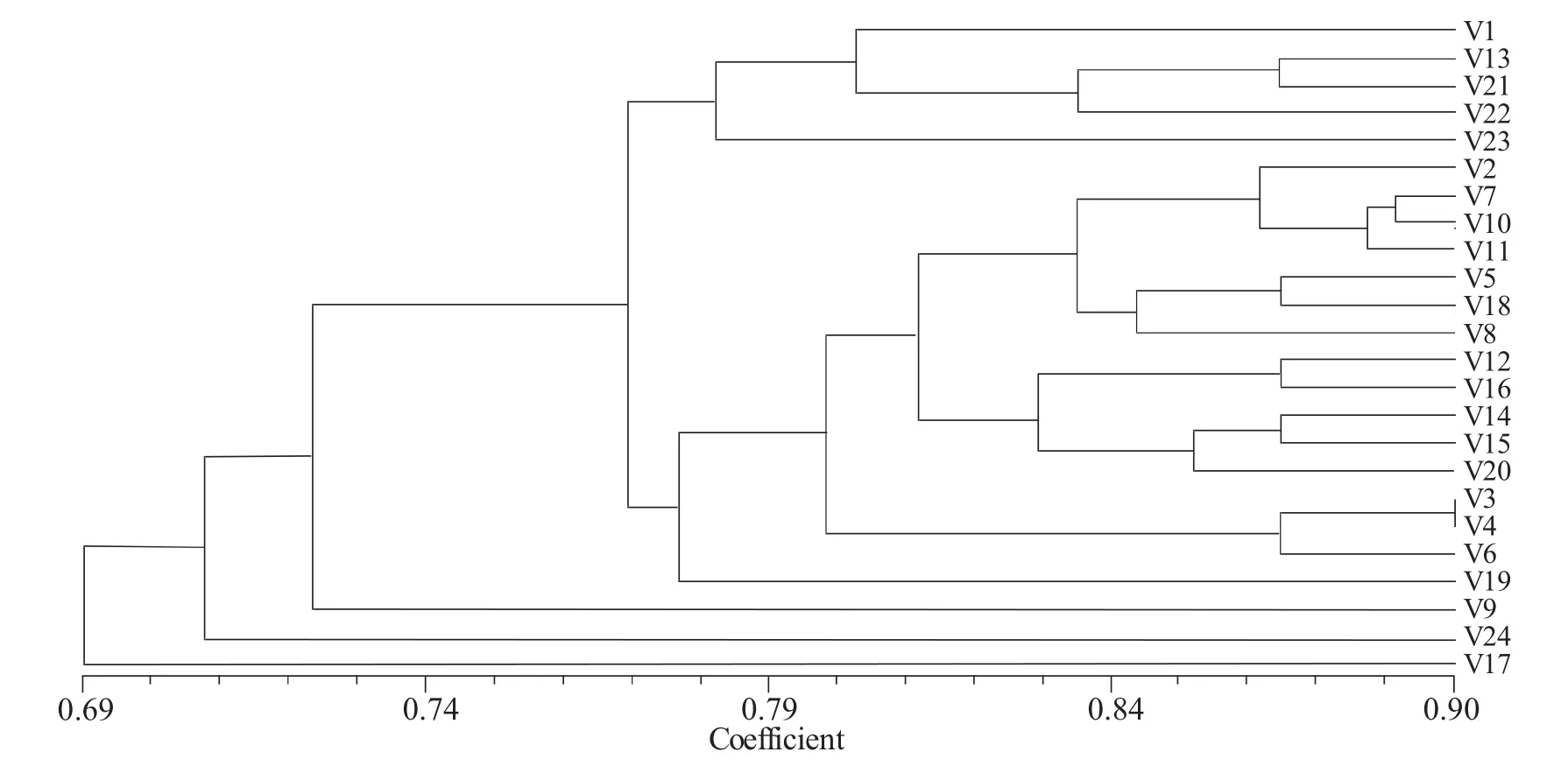
图4 24个亚麻栽培品种的聚类分析Fig.4 Clusteringdiagramfor 24 cultivars ofLinum usitatissmum L.
3 结论与讨论
3.1 在应用分子标记进行植物品种分类及亲缘关系鉴定过程中,引物的筛选是实验的关键,直接影响聚类结果。本实验中共筛选出的10个(5个RAPD和5个ISSR)引物扩增稳定、条带多、多态性丰富。利用这10个引物对包括多胚亚麻1-6Ha-3在内的24份材料(系)的基因组进行扩增,得到了115条清晰谱带,其中87条属于多态性标记,多态性位点的比例达75.7%。该结果很好地揭示了这24份资源的遗传多样性,由此鉴定出各材料间存在着遗传差异。根据这10个引物对每份材料的扩增结果,将供试的24份亚麻种质在阈值(相似系数)为0.73处分为四大类群,多胚品系1-6Ha-3独为第二类群,且与其他23个亚麻品种的遗传相似系数分布在0.6522~0.7739之间,说明该品系与其他材料间都存在着较大的遗传差异。并且从分子水平上鉴定出该多胚品系与黑亚10号(V24)、原 2006-13(V21)、黑亚 11 号(V19)、原 2005-15(V17)等品种的遗传距离较大,与原2005-21(V5)、98-338、SXY20等品种的遗传距离相对较近。一般地,遗传距离较远或不同类型的材料间杂交时杂交优势较高[15]。根据本试验结果,可在1-6Ha-3和与其遗传距离远的黑亚10号、原2006-13等品种间配制杂交组合,有可能从其杂交后代的多胚种子中获得杂交优势高的单倍体植株,采用化学加倍或自然加倍方法达到快速固定杂交优势的目的。因此,本实验结果可为利用多胚亚麻固定杂种优势的研究提供理论依据和指导。

表3 24个亚麻基因型相似性系数矩阵Table 3 The genotype similaritymatrixof24 flax
3.2 本研究通过RAPD和ISSR两种分子标记技术相结合的方法对认定多胚亚麻与其他栽培亚麻品种之间的关系的分析结果表明,该方法能较好的用于分析亚麻属种下样品的遗传距离,更能灵敏地揭示两个关系相近的个体之间的遗传差异。适合解决种下分类鉴定的问题。并能为扩大资源利用和研究提供新的线索。
3.3 由于在实验中发现一些品种(系)间虽然植物学形态特征存在着较大差异,但分子标记结果相似系数较高而聚为一类;也有部分品种(系)虽然形态表现出相似特征,但因分子标记结果相似系数小却未聚到一类。这可能是由于品种在长期种植、选择或杂交过程中,使其在地理分布、形态特征、生态类型等方面发生了较大的变异。因此,只有采用形态标记、细胞学标记及多种分子标记技术结合的方法,以更多不同基因型种质为试验样本群体,才能更全面准确的揭示遗传变异,为新品种选育制订策略,为亲本选配、后代遗传变异及杂种优势水平的预测提供预见性指导。为种质资源的利用、新品种(系)的选育提供科学依据。
[1]Lee WB,et al.application ofrandomamplified polymorphic DNA(RAPD)tosystematics ofsome species ofLiliumin Korea[J].Korean Journal ofPlant Taxonomy,1993,23(2):35-42.
[2]赵祥云,等.中国野生百合种质资源及其研究利用[M].北京:中国农业出版社,1994.
[3]张克中.百合种质扩繁、亲缘关系及雄性不育诱导研究[D].北京:北京林业大学,2003.
[4]赵庆芳.百合栽培品种资源的RAPD分析[J].兰州大学学报:自然科学版,2006,41(2):30-33.
[5]Wen C S,et al.Genetic differentiation of Lilium longiflorum Thunb.var.scabrum Masami in Taiwan using random amplified polymorphic DNAand morphological characters[J].Bot Bull Acad Sin,1999.40:65-71.
[6]左志锐,穆鼎.百合遗传多样性及亲缘关系的RAPD分析[J].园艺学报,2005,32(3):468-472.
[7]王振东,等.大豆抗旱种质资源遗传多样性的RAPD分析[J].大豆科学,2009,28(1):26-30.
[8]钟淮钦,等.观赏向日葵种质资源遗传多样性RAPD分析[J].分子植物育种,2009,7(1):73-78.
[9]XIE Fu-ti,et al.Phylogenetic analysis of vegetable-type(Edamame))and grain-type soybean [Glycinemax(L.)Merr.]cultivars through ISSR marker[J].Soybean Science,2008,27(5):732-739.
[10]HUO Guang,et al.Genetic diversityanalysis of 44 shares of hibiscus cannabinus L.germplasm resources using ISSR molecular[J].Agricultural Science&Technology,2009,10(3):63-67.
[11]王关林,方宏筠.植物基因工程原理与技术[M].北京:科学出版社,1998.
[12]黄文功.亚麻RAPD的反应体系优化及引物筛选[J].黑龙江农业科学,2009.2:4-6.
[13]丁明忠,等.用ISSR分析四川苎麻品种(系)间的遗传关系及雄性不育分子标记的建立 [J].核农学报,2008,22(2):183-187.
[14]汪恩华,等.形态与分子标记用于羊草种质鉴定与遗传评估的研究[J].草业学报,2002,11(4):68-75.
[15]别墅.中国3大主产棉区棉花品种遗传多样性的RAPD及其与农艺性状关系的研究[J].中国农业,2001,34(6):597-603.
