长白落叶松幼苗对铵态氮和硝态氮吸收的动力学特征
2010-10-08魏红旭徐程扬马履一江俐妮李雪莲杨卓
魏红旭,徐程扬*,马履一,江俐妮,李雪莲,杨卓
(1北京林业大学林学院,北京100083;2北京林业大学森林培育与保护重点实验室,北京100083;3吉林市龙潭区江密峰苗圃,吉林132206;4北京林业大学水土保持学院,北京100083)
随着农业生产集约化的发展,过量施肥特别是氮肥引起的诸多问题是植物营养、土壤、环境等学科关注的热点问题[1]。据报道,由于不合理施肥使所施氮肥的32%~85%无法被植物吸收利用[2-3]。除了氮素的淋失、挥发等损失外,植物对氮素的吸收、利用、转化以及根系的吸收动力学特征等因素都会影响氮肥的利用率[4]。
20世纪50年代初,Epstein和Hagen[5]首先将酶促反应动力学方程应用于植物对离子吸收的研究,开创了植物吸收养分动力学研究的先河。70年代以后,不断有学者对动力学方程进行改进,在这期间确立了经典的以米氏酶促反应动力学方程为基础的植物根系吸收离子的表达方程[6-7]。近年来国内外在养分吸收动力学研究方面已经开展了大量的工作,但所采用的试验材料多为水稻、大豆和生菜[8-12]等农作物,关于林业苗木的养分吸收动力学研究的报道相对较少[13-14]。
长白落叶松(Larix olgensis)是中国东北地区速生丰产林的主要树种,因其材质优良、速生丰产等特点,具有极强的经济价值和生态效益。生产上常规的长白落叶松养分管理方式十分粗放,不仅造成极大的人力和物力的浪费,而且不利于优质苗木的培育。本研究以当年生春季播种的长白落叶松幼苗为试验材料,在不同氮源供给的情况下探讨苗木对NH+4和NO-3吸收的动力学特征及其动态变化,以期为落叶松的动力学和养分培育方面的研究提供理论依据。
1 材料与方法
1.1 材料准备
试验材料为东北地区苗木生产普遍采用的当年生长白落叶松苗木,采集于吉林市龙潭区江密峰苗圃(126°45′E,43°45′N),其种源来自吉林省小北湖 。于2008年4月17日进行整地,以生产强度施入基肥后进行播种,播种量为5.5 g/m2,播种方法及出苗期水分管理与生产方式同步。分别于生长初期、速生期和木质化期分3次选取长势均一的苗木进行收获,每次收获量为150株,每次收获苗木生长情况见表1。将收获苗木带回实验室后洗净根系表面泥土,用去离子水饥饿处理24 h后用于养分动力学试验处理。
1.2 试验设计
不同氮素形态的吸收动力学试验采用离子耗竭法。设 3个营养液浓度处理:1)NH+4处理[以(NH4)2SO4为氮源];2)NO-3处理[以Ca(NO3)2为氮源];3)NH+4∶NO-3=1∶1 处理(浓度比,以NH4NO3为氮源,每种氮离子浓度与单一氮源时相同)。每处理设置 4 个浓度梯度:0.5、1.0、1.5、2.0 mmol/L,同时设置不供氮处理为对照(CK)。将经饥饿处理的苗木移入已装有130 mL营养液的瓶中,起始pH为6.5。为减少通气不均对根系吸收能力的影响,在每瓶中加入0.1 mL 3%的H2O2以提供O2。为防止NH+4硝化,在装有NH+4的营养液内加入7 μ mol/L的硝化抑制剂(C2H4N4)。每5株苗木为一个处理单位,每处理3次重复,以海绵固定,处理18 h。
1.3 测定方法
每次处理后取出苗木根系,切除地上部分,用吸水纸吸干根系表面水分后立即称重得到根系鲜重。测定处理后的营养液体积,同时测定其中NH+4、NO-3离子浓度。NO-3离子用直接比色法测定,NH+4离子采用纳氏试剂比色法测定[14],测定仪器为Agilent 8543型分光光度计。
1.4 参数计算方法
离子吸收速率[μ mol/(g◦h),plant]的计算参考杨洪强等[15]的方法:


表1 长白落叶松幼苗的生长情况Table 1 Growth parameters of Larix olyensis seedling under different growth periods
式中:C0、C1分别表示处理前后营养液的离子浓度(mmol/L);V0、V1分别表示处理前后营养液体积(L);T为处理时间(h);RFM为根系鲜重(g/plant,FM)。
Michaelis-Menten方程描述的计算采用双倒数Lineweaver-Burke法[16]:
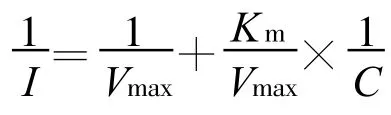
式中:I为离子吸收速率[μ mol/(g◦h),plant];C 为处理前离子浓度(mmol/L);Vmax为最大吸收速率[μ mol/(g◦h),plant];Km 为米氏常数。 Vmax反映了植物吸收某种离子的最大潜力,其值越大,表明吸收该离子的内在潜力就越大;同时Vmax值还与转运离子的载体数量和效率成正比[11]。Km为表观米氏常数,是1/2Vmax时的外液浓度,它表示载体植物根系对离子的亲和力,Km值越大,根系与养分离子的亲和力越小[13,17]。
辅助分析参数Cmin、α、β的计算分别参考华海霞等[16]和翟明普和蒋三乃[13]所报道的方法,具体如下:

式中:Cmin为离子吸收补偿点,表示净吸收速率为零时的营养液离子浓度,Cmin越小,根系的吸收能力越强[13];α值反映养分流入根系的速率,α值越大,养分离子流入根系的速率越快,反之越慢[13,15];β值是由前人提出的用来研究植物耐瘠薄能力的参数,其值的大小与植物耐瘠薄能力成反比[18]。
为了对比2种形态氮同时存在与单一形态氮存在间的动力学参数差异,引入一新参数Ii,其计算方法为:

式中:P2为NH+4和NO-3以1∶1浓度比同时存在时某一动力学参数值;P1为NH+4或NO-3单独存在时某一动力学参数值。两指标的Ii值差距越大,表明其中一种指标(较大者)对苗木的生理学影响越剧烈。
2 结果分析
2.1 单一氮源时不同生长期苗木对NH+4、NO-3的吸收动力学特征
单一氮源条件下,供试苗木在不同生长阶段对NH+4、NO-3的吸收速率随着NH+4、NO-3浓度的变化规律均符合Michaelis-Menten方程的描述,并且吸收NH+4、NO-3的速率(图1)和 Vmax、α值(表2)均随着苗木的生长呈下降趋势。说明随着苗木的生长吸收NH+4和NO-3的载体的数量和效率逐渐下降,导致离子流入根系的速率下降。生长初期和速生期,苗木对NH+4吸收的 Vmax、α值(表2)和速率均高于NO-3(图1),而在木质化期二者的 α值和吸收速率间均无明显差异。可见,单一氮源下苗木对于NH+4的吸收速率整体上强于NO-3。
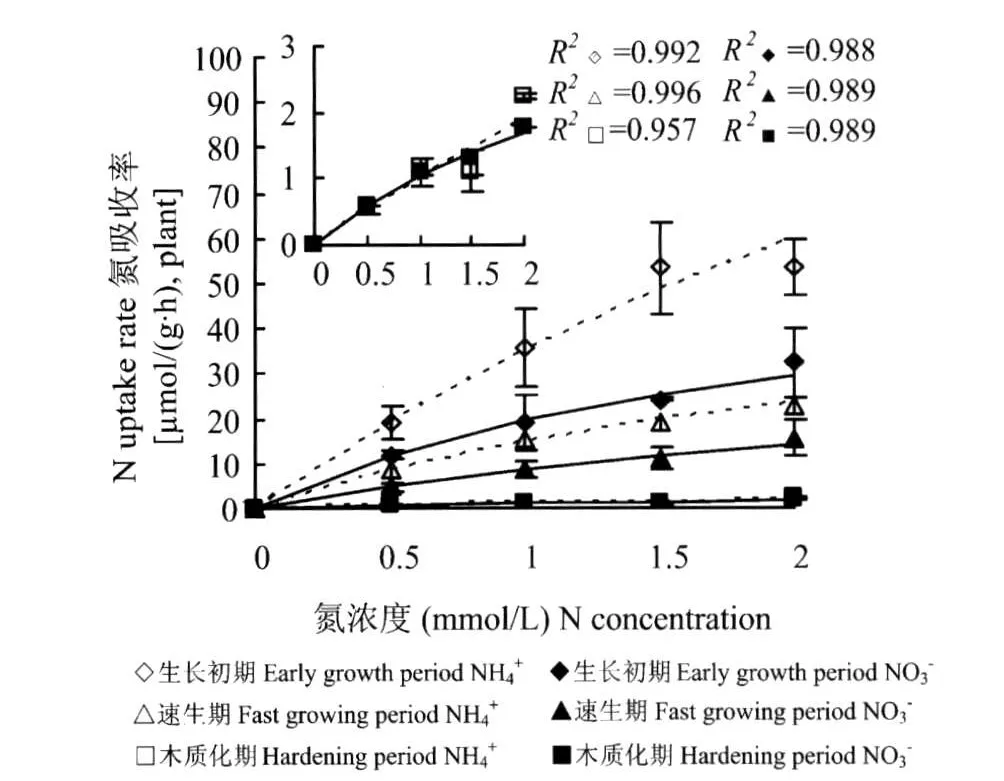
图1 不同生长期内单一(NH4)2SO4或Ca(NO-3)2供给下苗木对NH+4和NO-3的吸收动力学特征Fig.1 Kinetics of NH+4and NO-3uptakes by seedlings with NH+4or NO-3as N source separately in different growth periods
2.2 NH+4和NO-3共存时不同生长期苗木对NH+4的吸收动力学特征
无论NO-3存在与否,供试苗木在不同生长阶段对NH+4的吸收速率随着NH+4浓度的变化规律均符合Michaelis-Menten方程的描述(图2),并且NH+4的吸收速率均随着苗木的生长而下降。与无NO-3时相比,NO-3的加入大幅度降低了苗木在速生期对于NH+4的吸收速率,而并未明显影响苗木在生长初期和木质化期对于NH+4的吸收速率。
在不同生长阶段,苗木吸收NH+4的IiKm均大于IiVmax(表2),说明苗木对NH+4的吸收主要受载体亲和力的影响,而并非载体数量和效率。在生长初期和硬化期,IiKm和 IiVmax间差距分别只有4.738和0.001,并且加入NO-3后的Km较单NH+4时分别只提高了76.924和13.463。而速生期的IiKm和IiVmax间差距高达199.464,同时随着NO-3的加入,Km较单NH+4时提高了682.811。说明在速生期时苗木对于NH+4的吸收受亲和力的影响增加,NO-3的加入降低了根系对NH+4的亲和力,降低了NH+4流入根系的速率和吸收速率。而在生长初期和木质化期,由于根系对NH+4的亲和力较低,NO-3的加入并未对NH+4的吸收产生明显影响。
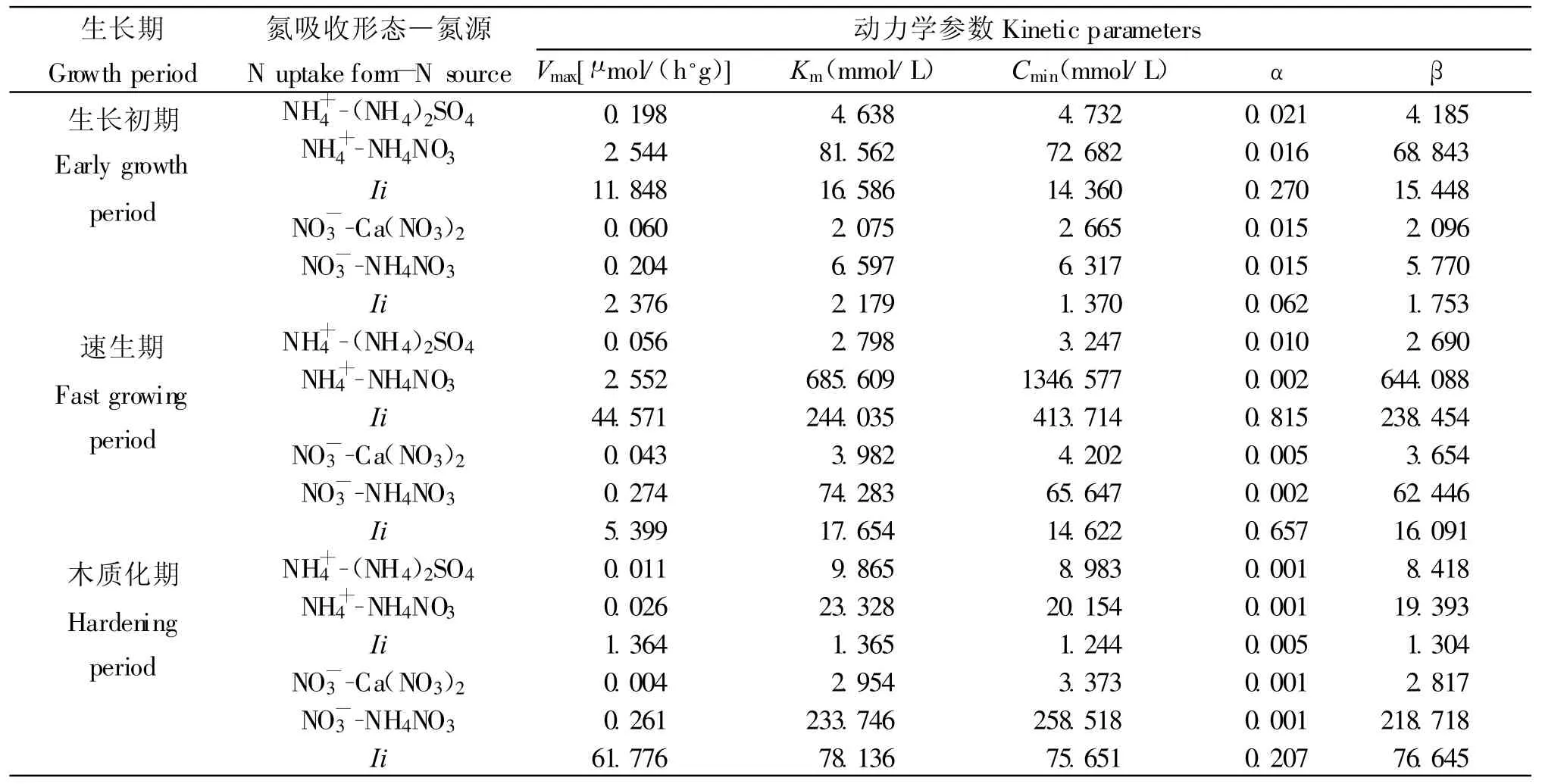
表2 不同生长期内不同氮源的NH+4、NO-3的吸收动力学参数动态Table 2 Dynamics of kinetic parameters of NH+4and NO-3uptakes from different N sources at different growth periods

图2 不同生长期内(NH4)2SO4或NH4NO-3供给下苗木对NH+4的吸收动力学特征Fig.2 Kinetics of NH+4uptake by seedlings with(NH4)2S O4or NH4NO3supplied in different growth periods
2.3 NH+4和NO-3共存时不同生长期苗木对NO-3的吸收动力学特征
无论NH+4存在与否,供试苗木材料在不同生长阶段对NO-3的吸收速率随着NO-3浓度的变化规律也同样显著符合Michaelis-Menten方程的描述,并且NO-3的吸收速率随着苗木的生长而下降(图3)。与单NO-3为氮源时相比,NH+4的加入(1∶1)降低了苗木在速生期对于NO-3的吸收速率,却大幅度提高了根系在生长初期对于NO-3的吸收速率,而对于木质化期的吸收促进的影响不明显。
长白落叶松苗木在生长初期对于NO-3的吸收主要受载体数量和效率影响(IiVmax=2.376;IiKm=2.179),虽然NH+4的加入使得Km提高了4.522(表2),NO-3的吸收速率仍较单NH+4为氮源时有所提高。进入速生期后,随着NH+4的加入NO-3流入根系的速率的有所下降(α=0.002),导致根系对NO-3的亲和力的影响强于载体数量和效率的影响(IiKm-IiVmax=12.255)。因此随着NH+4的加入,根系对NO-3的亲和力的下降导致NO-3的吸收下降。
3 讨论与结论

图3 不同生长期内Ca(NO3)2或 NH4NO3供给下苗木对NO3-的吸收动力学特征Fig.3 Kinetics of NO3-uptake by seedlings with Ca(NO3)2 or NH4NO3supplied in different growth periods
不同的植物种类在离子吸收上往往存在着较大的差异[10]。本次试验设置的 0~ 2.0 mmol/L的NH+4、NO-3营养液离子浓度在前人关于水稻[19]、生菜[11]和水曲柳[14]等试验材料的报道中都已被采用过,并证实其与根系吸收离子间的关系符合动力学描述。虽然本次试验结果亦发现了0~2.0 mmol/L的NH+4、NO-3营养液离子浓度与长白落叶松幼苗吸收速率间关系符合动力学描述(图1~图3),但是却没有出现前人发现的吸收速率曲线随着营养液离子浓度的增加而逐渐趋于平缓的规律。这也许是因为长白落叶松苗木对于NH+4、NO-3离子的吸收水平高于前人所采用的植物材料[11,19],也可能是由于前人所采用的材料的准备时间比较短[14],其具体原因有待进一步进行试验分析。
已有报道指出[19],增NO-3营养液可以促进根系对NH+4的吸收[8-9],但也有认为增加NO-3反而引起了NH+4的吸收速率下降[20]。本研究结果显示,在速生期,NO-3的加入抑制了载体与NH+4的亲和力,明显降低了NH+4流入根系的速度,从而导致速生期NH+4的吸收速率下降(图2)。但段英华等[19]则认为,NO-3对NH+4吸收的影响主要在于NH+4载体的运转速率而非吸收位点与NH+4之间的亲和性。前人的研究大多采用水稻等农作物为试验材料[8-9,19-20],得到的普遍结论是,NO-3的吸收会增加根系细胞内负电荷数量,有利于阳离子的积累[12]。但是,不可忽略的是,植物在吸收还原NO-3的时候需要消耗大量能量。苗木在速生期时由于叶片生长对于NH+4的需求较强[21],和NO-3的吸收还原造成的能量消耗,很有可能通过影响根系中谷氨酰胺合成酶(GS)的合成,进而影响NH+4的同化而降低亲和力,影响根系对其的吸收。而段英华等[19]试验发现,NO-3对于GS的活性具有促进作用,这可能与其测定GS的部位是叶片有关。由于缺少相关试验结果的支持,以上结论的合理性还需要通过以苗木为试验材料进一步进行试验验证。
有关NH+4影响NO-3的吸收的问题,目前很多研究结果比较统一,即NH+4的存在会明显抑制根系对NO-3的吸收[8-9,14,17]。王波等[11]将NH+4对NO-3吸收的影响途径归纳为2种:1)影响控制NO-3载体蛋白合成的基因表达,进而减少细胞膜上载体蛋白的总量;2)NH+4的存在可能影响载体所处细胞膜上的周边环境,例如膜极化程度的改变[22]、膜结构的改变[23]等。本研究结果显示,在速生期,NH+4的加入会抑制苗木根系与NO-3的亲和力(表2),进而影响NO-3的吸收速率(图3);但是在生长初期NH+4的加入会提高吸收NO-3的载体数量和效率,因此提高了根系对其的吸收速率。苗木在生长初期相对以根系生长为主,NH+4的加入有利于蛋白的合成,从而提高了吸收NO-3的载体数量,促进其吸收。此结果与王波等[11]报道的结果相反,可能与试验材料的种类和取样时期有关。在速生期时,苗木相对以地上部分生长为主,此时根系需要吸收大量的NO-3以满足地上部分生长的需要。有试验结果表明,根系在吸收NO-3的过程中要分泌大量的有机酸,降低根际周围溶液的pH[12]。任军等[14]认为在水曲柳的培养液中加入NH+4后影响NO-3吸收的原因是由于NH+4和H+的交换降低了根系周围溶液的pH值,使得根细胞液内OH-浓度升高,不利于NO-3的吸收。本次试验结果很可能是由于NH+4的加入降低了根际周围溶液的pH值,影响了有机酸的分泌,进而影响NO-3的吸收。但是,由于缺乏试验结果的支持,以上推断还有待进一步论证。
综上所述,长白落叶松苗木对于NH+4和NO-3的吸收速率随着生长而下降。当两种离子同时存在时,长白落叶松苗木在生长初期NH+4会提高根系吸收NO-3的载体数量和效率,从而提高其吸收速率;在速生期,两种离子会降低彼此与相应载体的亲和力,导致彼此的吸收速率都有所下降;在木质化期,由于苗木的生长基本已经停止,根系对于两种离子的吸收间没有明显差异。因此,在长白落叶松苗木的施肥措施中,应以铵态氮肥为主,如磷酸二铵、尿素和硫酸铵等,但是在生长初期可以适当增施硝态氮肥,如硝铵等,以提高苗木对氮素的利用效率。
[1] 王春阳,周建斌,郑险峰,等.不同栽培模式对小麦-玉米轮作体系土壤硝态氮残留的影响[J].植物营养与肥料学报,2007,13(6):991-997.Wang C Y,Zhou J B,Zheng X F et al.Effect of different cultivation methods on soil residual nitrate underwinterwheat-summermaize cropping systemp[J].Plant Nutr.Fert.Sci.,2007,13(6):991-997.
[2] Juntunen M L,Hammar T,Rikala R.Leaching of nitrogen and phosphorus during production of forest seedlings in containers[J].Environ.Qual,2002,31:1868-1874.
[3] Juntunen M L,HammarT,Rikala R.Nitrogen and phosphorus leaching and uptake by container birch seedlings(Betula pendula Roth)growth in three different fertilizations[J].New.For,2003,25:133-147.
[4] 孙敏,郭文善,朱新开,等.不同氮效率小麦品种苗期根系的NO3-、NH4+吸收动力学特征[J].麦类作物学报,2006,26(5):84-87.Sun M,Guo W S,Zhu X K et al.Kinetics of nitrate and ammonium uptake by different wheat genotypes at seedling stage[J].J.Trit.Crops,2006,26(5):84-87.
[5] Epstein E,Hagen C E.A kinetic study of the absorption of alkali cations by barley roots[J].Plant Physiol.,1952,27:457-474.
[6] Claassen N,Barber S A.A method for characterizing the relation betweennutrient concentration and flux into roots of intact plants[J].Plant Physiol.,1974,54:564-568.
[7] Baligar V C,Barber S A.Genotypic differences of corn for ion uptake[J].Agron.J.,1979,71:870-873.
[8] Kirk G J D.Plant-mediated processes to acquire nutrients:Nitrogen uptake by rice plants[J].Plant Soil,2001,232:129-134.
[9] Kronzucker H J,Siddiqi M Y,Class A D M et al.Nutrient ammonium synergism in rice:A subcellular flux analysis[J].Plant Plysiol.,1999,119:1041-1045.
[10] 孙小茗,封克,汪晓丽.K+高亲和转运系统吸收动力学特征及其受NH4+影响的研究[J].植物营养与肥料学报,2007,13(2):208-212.Sun X M,Feng K,Wang X L.Kinetics of high affinity system for K+and effects of NH4+[J].Plant Nutr.Fert.Sci.,2007,13(2):208-212.
[11] 王波,赖涛,沈其荣.不同铵硝配比营养液对典型生菜硝酸盐吸收动力学的影响[J].植物营养与肥料学报,2007,13(6):1098-1104.Wang B,Lai T,Shen Q R.Effects of NH4+-N/NO3--N ratios on kinetics of nitrate uptake by two typical lettuce genotypes in hydropoics[J].Plant Nutr.Fert.Sci.,2007,13(6):1098-1104.
[12] 高祖明,张春兰,倪金应,等.黄瓜等九种蔬菜与NO3--N亲和力的研究[J].南京农业大学学报,1990,13(1):75-79.Gao Z M,Zhang C L,Ni J Y et al.Study on compatibility of several vegerables with NO3--N[J].J.Nanjing Agric.Univ.,1990,13(1):75-79.
[13] 翟明普,蒋三乃.小钻杨和刺槐根系养分吸收的动力学研究[J].北京林业大学学报,2006,28(2):29-33.Zhai M P,Jiang S N.Dynamics of nutrient absorption in root systems of Populus×xiao zhuanica and Robinia pseudoacacia[J].J.Beijing For.Univ.,2006,28(2):29-33.
[14] 任军,徐程扬,林玉梅,等.水曲柳幼苗根系吸收不同形态氮的动力学特征[J].植物生理学通讯,2008,44(5):919-922.Ren J,Xu C Y,Lin Y M et al.Kinetic characteristics of different forms of absorbing nitrogen in root system of Fraxinus mandshurica Rupr.seedling[J].Plant Physiol.Commun.,2008,44(5):919-922.
[15] 杨洪强,张连忠,戚金亮,等.苹果砖木根系钙素吸收动力学研究[J].园艺学报,2003,30(3):253-257.Yang H Q,Zhang L Z,Qi J L et al.The kinetics of Calcium uptake in apple rootstock roots[J].Acta Hor.Sin.,2003,30(3):253-257.
[16] 华海霞,梁永超,娄运生,等.水稻硅吸收动力学参数固定方法的研究[J].植物营养与肥料学报,2006,12(3):358-362.Hua H X,Liang Y C,Lou Y S et al.Comparison of research methods for silicon uptake kinetics of rice[J].Plant Nutr.Fert.Sci.,2006,12(3):358-362.
[17] 赵越,马凤鸣,张多英.甜菜对不同氮素吸收动力学的研究[J].东北农业大学学报,2006,37(3):294-298.Zhao Y,Ma F M,Zhang D Y.Study on the absorption kinetics of different nitrogen in sugar beet[J].J.Northeast Agric.Univ.2006,37(3):294-298.
[18] 蒋廷惠,郑绍建,石锦琴,等.植物吸收养分动力学研究的几个问题[J].植物营养与肥料学报,1995,1(2):11-l7.Jiang T H,Zheng S J,Shi J Q et al.Several considerations in kinetics research on nutrients uptake by plants[J].Plant Nutr.Fert.Sci.,1995,1(2):11-17.
[19] 段英华,张亚丽,沈其荣.增硝营养对不同基因型水稻苗期吸铵和生长的影响[J].土壤学报,2005,42(2):260-266.Duan Y H,Zhang Y L,Shen Q R.Effect of nitrate on the ammonium uptake and growth of different genotypes of rice(Oryza sativa)at the seedling stage[J].Acta.Ped.Sin.,2005,42(2):260-266.
[20] 张亚丽,董园园,沈其荣,等.不同水稻品种对铵态氮和硝态氮吸收特性的研究[J].土壤学报,2004,41(6):918-924.Zhang Y L,Dong Y Y,Shen Q R.Characteristics of NH4+and NO3-uptake by rices of different genotypes[J].Acta.Pedol.Sin.,2004,41(6):918-924.
[21] 吴楚,王政权,范志强.氮素形态处理下水曲柳幼苗养分吸收利用与生长及养分分配与生物量分配的关系[J].生态学报,2005,25(6):1282-1291.Wu C,Wang Z Q,FanZ Q.Relationships between nutrient uptake,use efficiency and growth,and between nutrient partitioning patterns and biomass partitioning patterns in Fraxinus mandshurica seedlings supplied with different ratios of nitrogen forms[J].Chin.J.Appl.Ecol.,2005,25(6):1282-1291.
[22] Crawford N M,Glass A D M.Molecular and physiological aspects of nitrate uptake in plants(Reviews)[J].Trends Plants Sci.,1998,3(10):389-395.
[23] Colmer T D,Bloom A J.A comparision of NH4+and NO3-net fluxes along roots of rice and maize[J].Plant Cell Environ,1998,21:240-246.
