湖泊体系中长链烯酮研究进展
2010-09-07储国强刘国祥王晓华刘美美石丽明谢曼曼
孙 青, 储国强, 刘国祥, 王晓华, 刘美美,石丽明, 谢曼曼, 凌 媛
1)国家地质实验测试中心, 北京 100037;
2)中国科学院地质与地球物理研究所, 北京 100029;
3)中国科学院水生生物研究所, 湖北武汉 430072
湖泊体系中长链烯酮研究进展
孙 青1), 储国强2), 刘国祥3), 王晓华2), 刘美美1),石丽明1), 谢曼曼2), 凌 媛1)
1)国家地质实验测试中心, 北京 100037;
2)中国科学院地质与地球物理研究所, 北京 100029;
3)中国科学院水生生物研究所, 湖北武汉 430072
长链烯酮不饱和度(U37k′)作为定量反映古温度变化的重要替代指标, 已在海洋中得到广泛应用, 但在湖泊中长链烯酮不饱和度与温度的关系及其母源研究则很少。课题组研究了中国不同气候带、不同水化学环境湖泊表层沉积物中长链烯酮, 发现多数湖泊中存在2~4个不饱和键的长链烯酮, 首次报道硫酸盐型湖泊中存在长链烯酮, 总结了湖泊中长链烯酮的分布模式, 探讨了分布模式与环境、母源的关系。研究了湖泊长链烯酮不饱和度与温度的关系, 发现湖泊长链烯酮不饱和度与年均气温和春秋季节温度高度相关, 建立了中国湖泊表层沉积物中长链烯酮不饱和度与温度的经验函数关系, 结合文献中发表的数据, 建立了从热带的北缘湖光岩玛珥湖到北极的格陵兰的湖泊沉积物中长链烯酮不饱和度与温度的经验函数关系: U37k′= 0.031× T + 0.094 (n=76, r2= 0.67)。首次发现并成功分离出湖泊中长链烯酮母源等鞭金藻Chrysotila lamellosa,通过单藻种控温培养, 建立长链烯酮不饱和度与水温关系方程, 实验室培养公式与经验公式斜率一致, 验证了长链烯酮不饱和度温标, 表明长链烯酮是可靠的陆地温标。
湖泊; 长链烯酮; 不饱和度; 分布特征; 温度; 母源
古气候、古环境变化研究的关键问题之一是古气候、古环境变化定量指标的研究, 长链烯酮不饱和度温标, 由于不受碳酸盐溶解作用、沉积作用、氧化作用及长链烯酮丰度等因素的影响(Sikes et al., 1991; Prahl et al., 2003), 已成为继微体化石(有孔虫等)、氧同位素之后又一古气候变化研究的重要替代指标(孙青等, 2002)。尽管有研究表明营养、光照等环境因素可能影响长链烯酮不饱和度温标(Sikes et al., 2005), 但U37k′温标仍然广泛地、成功地应用于定量重建古海表水温(Rostek et al., 1993; Bard et al., 1997; Sachs et al., 1999; 垄庆杰等, 1999; Zhao et al., 2000; Kienast et al., 2001; Seki et al., 2004)。海洋环境中长链烯酮不饱和度温标(U37k′)不仅能够反映地质历史时期冰期-间冰期较大的温度变化, 而且能够定量反映中世纪暖期、小冰期及近代的温度波动(Abrantes et al. 2005)。
U37k′温标在全球海洋中得到广泛应用主要归功于: 1)长链烯酮在海洋中广泛存在(Müller et al., 1998); 2) 海洋中长链烯酮的母源比较清楚, 只有少数的几种金藻合成长链烯酮: 例如广海种Emiliania huxleyi、Gephyrocapsa oceanica(Volkman et al., 1980; Marlowe et al., 1984a, b)和滨海种Isochrysis galbana (Marlowe et al., 1984b; Conte et al., 1994; Versteegh et al., 2001)、Chrysotila lamellosa (Marlowe et al., 1984b, 1990; Rontani et al., 2004); 3)通过实验室对单藻种的控温培养以及全球海洋表层沉积物的研究,建立了U37k′-温度(T)的关系方程(Müller et al., 1998; Versteegh et al., 2001; Prahl et al., 1987; Volkman et al., 1995; Conte et al., 1995, 1998, 2001; Sawada et al., 1996; Rosell-Melé et al., 1995; Ternois et al., 1997; Pelejero et al., 1997; Bentaleb et al., 2002)。
长链烯酮不仅存在于海洋环境中, 而且还广泛存在于湖泊沉积物中(Cranwell et al., 1985; Li et al., 1996; 阳学贤等, 1996; Thiel et al., 1997; Innes et al., 1998; 盛国英等, 1998; Wang et al., 1998; Zink et al., 2001; 张干等, 2000; 孙青等, 2004; Chu et al., 2005)。湖泊地质历史时期沉积物中长链烯酮的研究发现Steisslingen湖的U37k′能够反映新仙女木事件前后的温度变化(Zink et al., 2001); 扎布耶盐湖的U37k′反映出末次冰期以来的温度变化(Wang et al., 1998);合同察汗湖U37k′值与δ18O有良好的对应关系, 可能反映出小冰期的降温事件(张干等, 2000); Titicaca湖(Theissen et al., 2005)的研究结果表明如果长链烯酮的母源能够确定, 长链烯酮不饱和度温标有可能成为湖泊古气候研究的定量指标。但湖泊中U37k′温标仍然处于探索中, U37k′能否成为定量重建湖泊古温度变化的重要替代指标,有待进一步系统研究湖泊体系中长链烯酮的分布、母源特征, 通过母源的培养和湖泊沉积物的研究获得可以被接受的U37k′-T关系方程。
1 湖泊中的长链烯酮分布特征
长链烯酮广泛存在于淡水湖(Cranwell et al., 1985; Zink et al., 2001; Chu et al., 2005))、咸水湖(Li et al., 1996; Wang et al., 1998; Abrantes et al., 2005)及碱性咸水湖(Thiel et al., 1997; 盛国英等, 1998;张干等, 2000)的湖泊沉积物中。我们首次报道了硫酸盐型盐湖中存在长链烯酮(Sun et al., 2004), 比较系统地研究了中国不同气候带不同水化学类型的湖泊表层沉积物中的长链烯酮分布特征, 这些湖泊盐度变化很大, 从淡水湖到盐湖, 湖泊水化学类型包括碳酸盐型、硫酸盐型和氯化物型, 长链烯酮的含量范围 0.03-39.00(mg/g 干样), 其中碳酸盐型咸水湖夏日淖尔湖的沉积物中长链烯酮浓度最高, 硫酸盐型盐湖沉积物中长链烯酮的浓度最低(Sun et al., 2004, Chu et al., 2005)。
1.1 C37:4优势模式
中国的湖泊表层沉积物中长链烯酮C37:4含量(C37:4含量占总C37长链烯酮含量的百分比)变化范围是5%~96%,平均值55%(Chu et al., 2005; Sun et al., 2004; Liu et al., 2006; Liu et al. 2008), 在四海龙湾玛珥湖 1600年的沉积物(王晓华, 2009)中C37:4含量介于57.45%~39.95%, 平均值为46.86%, 青海湖3500年来沉积物(Liu et al., 2006)中C37:4含量介于: 15%~45%, 与世界其它地区湖泊沉积物中C37:4含量可比, 例如, Titicaca湖沉积物(Theissen et al., 2005)中C37:4含量为 8%~18%, 平均 12.5%, 德国的Steisslingen湖(Zink et al., 2001)为21%~66%, 土耳其的Van湖(Thiel et al., 1997)为20%~72%, 美国的北部平原区湖泊(Toney, Huang et al. 2010)C37:4含量为62%~69%; 湖泊中分离出的C. lamellosa的培养产物中检测出C37:4含量为6.3%~24.9%(Sun et al., 2007),分布在湖泊沉积物中长链烯酮C37:4含量变化范围之内。滨海环境沉积物和物种C. lamellosa和I. galbana的培养产物中检测出高含量C37:4(Rontani et al., 2004; Marlowe et al., 1984)。在开阔海洋沉积物和物种Emiliania huxleyi、Gephyrocapsa oceanica培养物中C37:4含量很低, 相对高的C37:4含量局限于低温地区(Schulz et al., 2000, Mercer et al., 2005, Sikes et al., 1991; Sikes and Volkman, 1993; Rosell-Mel et al., 2002; Harada et al., 2003)。虽然在美国东北部桑德希尔斯湖区(Toney et al., 2010)发现一些不含C37:4的湖泊, 但总体上, 滨海和内陆湖泊的沉积物中和母源合成的C37:4含量远比开阔海洋体系高得多, C37:4优势模式可能是湖泊系统和滨海环境中长链烯酮的普遍分布特征(图1。Sun et al., 2004; Chu et al., 2005; Cranwell 1985; Li et al., 1996; Zink et al., 2001)。
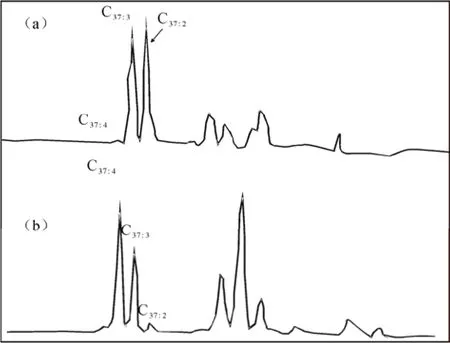
图1 湖泊和海洋沉积物中长链烯酮的分布特征a-非C37:4优势模式; b-C37:4优势模式Fig. 1 Distribution of long-chain alkenones in lake and marine sediments a-non-C37:4dominant pattern; b-C37: 4 dominant pattern
早期的研究表明, C37:4的百分含量可能受控于温度(Prahl et al., 1988)。在较冷的海洋环境中, 当C37:4含量的相对含量低于 5%时, C37:4含量与上覆海水温度存在很好的线性关系(Rosell-Mel et al.,1998),随后的研究表明, 在海洋环境中C37:4的含量与温度之间不存在线性关系的(Prahl et al., 1988; Freeman and Wakeham, 1992)。但我们对湖泊种C. lamellosa的培养发现, 在其他环境条件不变的情况下, 当温度从 22℃降到 10℃时, C37:4由 6.3%增长到 24.9% (Sun et al., 2007), 我们的结果支持温度是引起C37:4含量变化的重要因素的观点。
一些学者认为C37:4含量可能作为一种古盐度的替代指标(Schulz et al., 2000; Rosell-Mel et al., 1998, 2002), Ace湖(Coolen et al., 2004)、青海湖(Liu et al., 2008)中长链烯酮的C37:4与盐度之间具有一定的关系; C37:4可能记录了青海湖晚全新世湖水盐度变化(Liu et al., 2006)。但是, 无论湖泊(Chu et al., 2005)、还是海洋环境(Rosell-Mel et al., 1998; Lopez et al., 2005; Sikes and Sicre, 2002; Mercer et al., 2004)的研究结果都表明C37:4甲基酮与盐度之间的关系还存在不确定性, 目前, 没有一个全球性的C37:4和盐度关系。我们认为在复杂的湖泊体系中, 母源、温度、营养物质、盐度等生物、环境因子多变, 在某一地区或某一时段内C37:4含量变化可能与盐度相关, C37:4与盐度的关系可能不单纯是定鞭金藻的单一 属种对盐度变化的生理响应, 而是多种合成长链烯酮的藻类为适应盐度的变化发生种属间相对丰度 变化(Coolen et al., 2004), 从而导致长链烯酮的C37:4变化。
1.2 C37优势模式和C38优势模式
湖泊沉积物中的 C37/C38变化范围很大(0.02-7.8)(chu et al., 2005), 按链长特征, 湖泊中长链烯酮分布模式有 C37优势模式(C37>C38)和 C38优势模式(图2, C38> C37)。

图2 湖泊中长链烯酮分布模式: C38优势模式(a)和C37优势模式(b)Fig. 2 Distribution of long-chain alkenones in lake: C38 dominant pattern (a) and C37 dominant pattern (b)
C37优势模式是湖泊环境(Cranwell,1985; Li et al., 1996; Thiel et al., 1997; Wang and Zheng, 1998; Zink et al., 2001)、边缘海环境(Freeman and Wakeham, 1992; Schulz et al., 2000)和开阔海洋环境中(Prahl et al., 1988; Sikes and Volkman, 1993)最典型的模式。在开阔海洋环境的沉积物和物种中C37/C38较低, 多小于2.0(Chu et al., 2005), 而在湖泊沉积物和边缘海中的C37/C38的比值不乏高值的报道。无论是在中国湖区(Sun et al., 2004; Chu et al., 2005; Liu et al., 2006; Liu et al., 2008)、英国的湖区(Cranwell, 1985)、美国的东北桑德希尔斯和北部大平原区的湖泊(Toney et al., 2010)、Van湖(Thiel et al., 1997), 还是波罗的海 (Schulz et al., 2000) 的沉积物中都有高C37/C38比值的报道。相应的滨海和湖泊环境长链烯酮的母源C .lamellosa和I. galbana合成的长链烯酮的 C37/C38变化范围也较大, C. lamellosa从1.4到9.5(Rontani et al., 2004; Marlowe et al., 1984; Prahl et al., 1988; Sun, 2007), I. galbana从5.4到15.1(Marlowe et al., 1984)。滨海和湖泊的沉积物、相应环境中母源物种合成的长链烯酮的C37/C38烯酮比值整体上高于开阔海洋沉积物和长链烯酮的母源合成的C37/C38烯酮比值, 但是大多数情况下, C37/C38 的变化由藻类细胞的营养、生理和生长温度贡献 (Conte et al., 1998), C37/C38很难区分物种, 只有当C37/C38很高时, 可能合成长链烯酮藻类是等鞭金藻 C. lamellosa和I. galbana而非颗石藻 Emiliania huxleyi和 Gephyrocapsa oceanica。
C38优势模式在少数几个边缘海(Cacho et al., 1999; Schulz et al., 2000)、少数湖泊沉积物(Thiel et al., 1997; Innes et al., 1998; Zink et al., 2001)中出现。C38优势模式在硫酸钠或硫酸钠镁型湖泊沉积物中出现的几率高于淡水湖、碳酸盐型和硫酸镁型盐湖,例如在西班牙的硫酸钠或硫酸钠镁型湖泊广泛存在C38优势模式, 在中国的青海的大柴旦、小柴旦、内蒙古的莲花池等硫酸钠或硫酸钠镁型湖泊沉积物中长链烯酮的分布模式也多为C38优势模式。
1.3 C38乙基酮模式和C38甲基乙基酮模式
依据 C38长链烯酮的组成特征, 湖泊中的长链烯酮可以分为C38乙基酮模式和C38甲基乙基酮模式(图3)。C38乙基酮模式是指C38烯酮由C38乙基酮组成, C38甲基乙基酮模式是指 C38烯酮由C38乙基酮和C38甲基酮组成。综合研究不同母源藻种培养产物中长链系统分布特征(Volkman et al., 1980; Marlowe, Brassell et al. 1984; Rontani et al., 2004; Chu et al., 2005; Sun et al., 2007), 认为E. Huxleyi、G. Oceanic和个别Isochrysis galbana种属合成的长链烯酮为C38甲基乙基酮模式(Chu et al., 2005), 而C. lamellosa单藻种培养产物中未检出C38甲基酮,为C38乙基酮模式(Sun et al., 2007)。中国的巴里坤湖沉积物中 C38长链烯酮为甲基乙基酮模式, 而夏日淖尔湖沉积物中长链烯酮为 C38乙基酮模式, 与之相对应, 巴里坤湖沉积物中存在微体古生物化石G. oceanic(Chu et al., 2005), 夏日淖尔湖分离出了合成长链烯酮的母源C. lamellosa(Sun et al., 2007), 表明利用C38乙基酮模式和C38甲基乙基酮模式可以区分长链烯酮的母源。但是C38长链烯酮分布模式并没有得到广泛的重视,在湖泊长链烯酮研究的文献中只有部分文献给出了 C38长链烯酮的组成特征。如英国湖区Coniston湖(Cranwell, 1985)不同深度的沉积物中 C38长链烯酮分布模式不同, 在90~100 cm 为乙基酮模式, 而在 126~145 cm 和153~165 cm的沉积物中为甲基乙基酮模式, 中国的巴里坤湖(Chu et al., 2005)、南美的Titicaca(Theissen et al., 2005)、美国的Crose Mere湖沉积物和Upton Broad湖沉积物(Toney et al., 2010)、南极的Fryxell湖(Jaraula et al., 2010)、格陵兰的湖泊(D'Andrea et al., 2005)、美国的Brush湖和德国的Steisslingen湖(Zink et al.,2001)沉积物中为甲基乙基酮模式。中国的扎布耶湖(Wang et al., 1998)、俄罗斯的Pichozero湖(Zink et al., 2001)、英国Windermere湖(Cranwell, 1985)、南极的Ace湖(Coolen., 2004)、中国的青海湖的C38长链烯酮分布模式为乙基酮模式(Li.,1996; Liu et al., 2008)。如何利用长链烯酮的分布特征, 研究长链烯酮的母源可能是将来的研究内容。
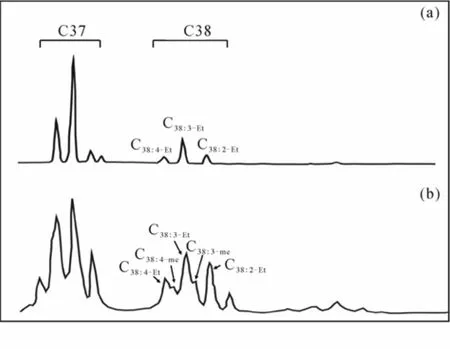
图3 C38长链烯酮的组成特征: C38乙基酮模式(a)和C38甲基乙基酮模式(b)Fig. 3 Composition of C38 long-chain alkenones: C38 ethylketone pattern (a) and C38 methylethylketone pattern (b)
2 湖泊中长链烯酮的母源
自20世纪80年代发现湖泊沉积物中存在长链烯酮以来, 学术界一直不断地探索湖泊沉积物中长链烯酮母源。但是, 由于湖泊长链烯酮母源藻类丰度低、个体小, 分离母源难度较大, 学术界一直未能解决此问题。Cranwell等(1985)认为单鞭金藻可能是湖泊沉积物中长链烯酮的主要来源。Zink等(2001)根据湖泊水化学特征和长链烯酮的分布特征, 认为湖水的酸碱度控制了产生长链烯酮的藻类的生长速度, 研究不同深度的水样发现叶绿素含量最高处的水样和大于 10μm的过滤样中存在长链烯酮, 认为长链烯酮来自大于10μm的藻类, 并通过藻类培养、长链烯酮和藻类分布的研究明确排除了湖泊中许多藻类是合成长链烯酮的母源, 例如: 与海洋定鞭藻纲对应的淡水物种 Chrysochromulina polylepsis 等,其他的还有绿藻门的 Phacotus lenticularis, 蓝藻门的 Synechocystis sp. 等, 亲碱性的硅藻 Cyclotella meneghiniana等, 但未能发现长链烯酮的母源, 说明长链烯酮可能并非来源于湖泊中的优势藻类, Zink等推测Chrychromulina parva有可能是某些湖泊中长链烯酮的母源。在我们研究的淡水湖中, 四海龙湾玛珥湖沉积物中长链烯酮含量最高, 而且沉积物中存在高丰度的金藻孢子, 推测淡水湖泊中长链烯酮的母源可能为金藻(Chu et al., 2005), 但藻类培养未能获得四海龙湾玛珥湖中合成长链烯酮的母源。
在中国内陆一些咸化湖, 如巴里坤湖、艾比湖表层沉积物中存在 Gephyrocapsa oceanica、Coccolithus pelagicus等颗石藻化石(徐钰林和孙镇城, 1998; 孙镇城等, 1997; 孙镇城等, 2002)。Chu等(2005)根据中国内陆一些咸化湖泊表层沉积物存在Gephyrocapsa oceanica等颗石藻化石、在相应 的湖泊沉积物中检出长链烯酮, 以及巴里坤湖长 链烯酮的分布模式与颗石藻G. oceanica培养产物的分布模式一致, 认为一些颗石藻(G. oceanica, C. pelagicus)可能是某些湖泊中长链烯酮的母源。在咸化湖泊中, 我们选择了长链烯酮含量高的夏日淖尔湖(碳酸盐型盐湖), 采集藻种, 培养分离, 在夏日淖尔湖采集的春秋季节的藻种培养样中, 我们(Sun et al., 2007)发现并成功分离了Chrysotila lamellosa, 单藻种培养表明Chrysotila lamellosa在生长过程中合成长链烯酮。
不同类型湖泊、同一湖泊不同发展阶段的沉积物样品中长链烯酮分布特征存在差异, 考虑到湖水物理、化学环境的巨大差异, 以及前人在长链烯酮母源等方面的研究结果, 我们认为湖泊中长链烯酮的母源不仅是 Gephyrocapsa oceanica、Chrysotila lamellosa, 可能还存在其他母源。近年来, 一些学者结合长链烯酮的分布特征和 18S rDNA序列, 认为Ace湖和 Fryxell湖中长链烯酮的可能来源是Isochrysis sp.和 C. lamellosa, 格陵兰不同湖泊中长链烯酮的母源不同, 一些可能来源于 Isochrysis sp.和 C. lamellosa(Coolen et al., 2004; Jaraula et al., 2010), 而另外一些湖泊, 例如美国的东北桑德希尔湖区的湖泊沉积物中不含 C37:4长链烯酮, 与其它湖泊截然不同的烯酮分布模式可能表明这些湖泊中长链烯酮的母源不是Isochrysis sp.和C. lamellose.而是其它物种。
3 长链烯酮不饱和度与温度的关系
孙青等(2004)报道了硫酸盐型盐湖中U37k′与湖区年平均温度存在较好的相关性, 硫酸盐型盐湖的U37k′温度曲线方程是U37k′=0.028× T +0.166(r2=0.88),此后, 我们(Chu et al., 2005)比较系统的总结了中国不同气候带不同水化学成分的 50个湖泊表层沉积物中的长链烯酮数据, 探讨了U37k′与年均气温、春夏秋冬和7月平均气温间的关系, U37k′与年均气温相关性最好U37k′= 0.0328 × T + 0.126(r2=0.83), 方程的斜率(0.0328)与实验室培养的E. huxleyi(0.033, Prahl and Wakeham, 1987)、世界范围内沉积物的标定(0.033, Müller et al., 1998)、南中国海(0.031, Pelejero and Grimalt, 1997)、圣芭芭拉盆地(0.034, Zhao et al., 2000和南海(0.038, Sikes et al.,1997) 的U37k′-T方程斜率一致(Chu et al., 2005), 结合近年来发表的北极格陵兰的湖泊(D'Andrea et al., 2005)、中国青海湖(Liu et al., 2008)、柴达木盆地湖泊(付明义等, 2008)、德国的湖泊(Zink et al., 2001)和美国北部大平原湖区(Toney et al., 2010)湖泊沉积物中获得的长链烯酮数据, 获得新的湖泊体系长链烯酮不饱和度与温度的关系: U37k′= 0.031× T + 0.094 (n=76, r2= 0.67)(图4)。次相关的是U37k’与春秋季节温度间的关系, 与夏冬季节均温相关性最差(Chu et al., 2005),可能表明长链烯酮的母源在春秋季节繁盛, 在此期间合成长链烯酮, 并记录了此期间的温度。湖泊可能的长链烯酮的母源是金藻 (Cranwell, 1985; Versteegh et al., 2001), 它们是典型的浮游植物, 以季节性生长为特征(Duff et al., 1995), 生长季节是早春和晚秋, 采集夏日淖尔湖藻类样品, 在4、5、9、10月的藻种培养样中发现了合成长链烯酮的藻类Chrysotila lamellosa, 而夏季藻类样品的混合培养样中虽然含有痕量的长链烯酮, 但没有发现Chrysotila lamellosa, 也可能说明长链烯酮不是在 7月合成的,所以U37k′与春、秋季节温度相关性优于其它季节是合理的, 不同湖泊的季节性藻类繁盛可能是削弱U37k′温度相关性的重要因素 (Prahl et al., 2001; Bac et al., 2003)。
在温度较高和温度较低的区域, U37k′与温度之间有明显的偏离(Sikes and Volkman, 1993; Rosell-Mel et al., 1995; Volkman et al., 1995; Conte et al., 1998; Conte et al., 2001; Versteegh et al., 2001; Mercer et al., 2005)。湖泊沉积物U37k′与温度间关系的标定方程在温暖端斜率有变缓趋势(Chu et al., 2005), 湖泊中长链烯酮的母源C. Lamellosa的温控培养实验结果表明在低于 14℃时, U37k′与温度关系方程的斜率也是变小的(Sun et al., 2007)。这种U37k′与温度间的关系方程在高、低温端斜率变小现象在海洋沉积物研究和单藻种培养研究中是一种普遍的现象(Sikes and Volkman, 1993; Sonzogni et al., 1997; Conte et al.1998)。可能说明合成长链烯酮的藻类细胞在适宜生长温度范围之外, 为适应温度变化调整长链烯酮相对含量的能力下降。

图4 湖泊体系长链烯酮不饱和度与温度的关系Fig. 4 Relationship between unsaturation and temperature of long-chain alkenones in lake system
以前的研究表明海洋体系U37k′-T间的关系可能受物种因素影响较小(Müller et al.,1998)。在海洋体系中, 一般的沉积物标定中U37k′-T具有高的相关系数, 海洋体系中物理化学和生物条件相对均一, 而且沉积物的标定的优点在于包含了环境变化、物种的分布变化、成岩作用的影响, 沉积物中长链烯酮的信息中短期的和合成环境中非温度的因素影响被平均、被削弱(Müller et al., 1998; Sachs et al., 1999, Conte et al., 2001), 然而, 一些研究表明基因的差异和生理响应的差异确实存在 (Volkman et al., 1995;Versteegh et al., 2001), 而且湖泊体系的水化学、生物物种、季节性变化远远大于海洋体系的变化, 尤其水化学的巨大差异可能导致湖泊体系生物物种发生重要改变, 考虑到湖泊中长链烯酮分布模式的不同, 在研究的湖泊中可能存在生态迥异、对环境尤其是温度变化响应具有很大差异的合成长链烯酮的物种, 考虑到当湖水盐度≤3.0 g/L, 生物区系没有明显差别(Hammer, 1986; Beadle, 1959), 我们将湖泊体系分为两组: 淡水-微咸水(盐度≤_3.0 g/L)和咸水湖。淡水-微咸水湖U37k’与年均大气温度的关系是U37k′= 0.037× MAAT + 0.108 (n=14, r2= 0.90, Chu et al., 2005), 盐湖的U37k′与年均大气温度的关系是U37k′= 0.025× MAAT + 0.153 (n=24, r2= 0.67, Chu et al., 2005)。淡水-微咸水湖的回归方程的斜率与咸水湖的差别很大, 斜率的不同可能是因为淡水-微咸水和咸水湖生物区系发生明显的变化, 合成长链烯酮的藻类可能有很大的差异。咸水湖回归方程的斜率与中国硫酸盐型湖泊U37k′-T 关系方程的斜率(Sun et al, 2004)相似, 与夏日淖尔湖中分离出长链烯酮母源(Chrysotila lamellosa)单藻种控温培养获得的线性回归方程: U37k′= 0.0257× T -0.2608 (n = 9, r2= 0.97, Sun et al., 2007)相似, 但与滨海种C. lamellosa的U37k′-T方程斜率相差很大(0.01, Rontani et al., 2004), 斜率的差异可能是由于不同种属之间对温度响应的变化不同造成。种属间遗传型变化可能是影响生物合成长链烯酮对温度的响应方式的重要因素(Conte et al., 1995; Yamamoto et al., 2000)。这个结果表明在某一湖泊中分布模式不同或奇特时, 利用U37k′重建湖泊古温度需要谨慎。
在培养实验中, C37:4含量从 10°C的 24.9%变至22°C的 6.3%, 表明C37:4在盐度不变的情况下随温度而变化(Sun et al., 2007)。当考虑到指标中引入C37:4,利用U37k指数(Brassell et al. 1986), 我们可以得到C. lamellosa培 养 产 物 的 U37k与 T线 性 关 系 : U37k=0.0377× T – 0.5992(n=14, r2=0.98, Sun et al, 2007), 显然生长环境的水温控制着U37k, 与海洋物种E. huxleyi和 G. oceanica 的培养结果一致(Conte et al., 1998, Conte et al., 1994); 德国的淡水湖沉积物中U37k与湖水温度呈线性关系(Zink et al., 2001)。但是C37:4的影响因素较多, 例如可能反映盐度(Rosell-Mele′et al., 1998; Rosell-Mele′et al., 2002; Liu et al., 2006), 虽然C37:4与盐度的关系不确定(Chu et al., 2005, Lopez et al., 2005), 而且C37:4较C37:2和C37:3易于氧化, 因此U37k′指标可能比U37k可靠(Sun et al., 2007)。最近, Pearson(2008)在研究西班牙盐湖中长链烯酮时提出了基于C37: 4-2和C38: 4-2长链烯酮同系物的新的温度指标, 西班牙不同盐湖中 U3K738与温度的关系是: U3K738= 0.0464×MAutAT- 0.867 (r2=0.80, n=13)。实验室培养和湖泊表层沉积物中长链烯酮温标的标定方面的研究促进了湖泊古温度计——长链烯酮不饱和度温标的定量化研究。
4 结论
长链烯酮不饱和度(U37k′)作为定量反映古温度变化的重要替代指标, 已在海洋中得到广泛应用,但在湖泊中长链烯酮不饱和度与温度的关系及其母源研究则很少。我们课题组研究了中国不同气候带、不同水化学环境湖泊表层沉积物中长链烯酮, 发现多数湖泊中存在 2~4个不饱和键的长链烯酮, 认为不同的长链烯酮分布模式可能与不同的环境和物种相关。首次报道硫酸盐型湖泊中存在长链烯酮。研究了湖泊长链烯酮不饱和度与温度的关系, 发现湖泊长链烯酮不饱和度与年均气温和春秋季节温度高度相关, 结合近年来不同研究者发表的不同地区湖泊中长链烯酮数据, 建立了湖泊表层沉积物中长链烯酮不饱和度与温度的经验函数关系。考虑到湖泊水化学的巨大差异可能导致湖泊体系生物物种发生重要改变, 我们将湖泊体系分为两组, 分别建立了两组不同水化学成分的湖泊: 淡水-微咸水和咸水湖中长链烯酮不饱和度与温度的经验函数。首次发现并成功分离出湖泊中长链烯酮母源等鞭金藻Chrysotila lamellosa, 通过单藻种控温培养, 建立长链烯酮不饱和度与水温关系方程, 实验室培养公式与经验公式斜率一致, 验证了长链烯酮不饱和度温标, 研究表明湖泊环境和海洋环境中长链烯酮的母源合成烯酮的路径和对温度响应的机制一致, 长链烯酮有可能成为可靠的陆地温标, 在陆地古气候变化研究中将发挥重要的作用。但是湖泊体系影响因素多, 相对海洋体系复杂, 湖泊长链烯酮温标的选择与应用需要谨慎, 长链烯酮温标的野外标定和母源分离培养等定量研究有待进一步加强。
付明义, 刘卫国, 李祥忠,徐黎明,王政,安芷生. 2008. 青海湖及柴达木盆地地区现代湖泊沉积物中长链烯酮的分布特征[J]. 湖泊科学, 20(3): 285-290.
刘传联, 赵泉鸿, 汪品先. 2002. 从化石群及壳体同位素看古近纪东营湖湖水化学[J]. 地球学报, 23: 237-242.
垄庆杰, 吴良基, 吴时国, 罗又郎, .张干. 1999. 南海长链烯酮化合物的检测及U值的应用[J]. 地球化学, 28 (1): 51- 57.
盛国英, 蔡克勤, 阳学贤, 卢家烂, 贾国东, 傅家谟, 彭平安. 1998. 合同察汗淖(碱)湖沉积物中长链不饱和烯酮及其古气候意义[J]. 科学通报, 43 (10): 1090-1094.
时兴合, 赵燕宁, 戴升, 徐亮, 李应业, 贾红莉, 张青梅. 2005.柴达木盆地40多年来的气候变化研究[J]. 中国沙漠, 25(1): 123-128.
孙青, 储国强, 李圣强, 吕彩芬, 郑绵平. 2004. 硫酸盐型盐湖中的长链烯酮及古环境意义[J]. 科学通报, 49: 1789-1792.
孙青, 储国强. 2002. 长链烯酮不饱和度温标研究进展[J]. 地质地球化学, 30(4): 63-67.
孙镇城, 杨藩, 张枝焕, 李守军, 李东明, 彭立才, 曾学鲁, 徐钰林, 茅绍智, 王强. 1997. 中国新生代咸化湖泊沉积环境与油气生成[M]. 北京: 石油工业出版社.
孙镇城, 杨革联, 乔子真, 杨藩, 李东明, 景民昌. 2002. 我国咸化湖泊沉积中钙质超微化石特征及其地质意义[J]. 古地理学报, 4: 56-63.
王晓华. 2009. 东北四海龙湾玛珥湖 1500年来长链稀酮不饱和度温标与古水温重建[D]. 北京: 中国科学院地址与地球物理研究所.
徐钰林, 孙镇城. 1998. 中国西北地区第四纪盐湖沉积中钙质超微化石的发现及其古环境意义[J]. 现代地质, 12: 49-55.
阳学贤, 盛国英, 卢家烂, 傅家谟. 1996. 合同查汗淖(碱)湖沉积物中的生物标志物特征及古环境意义[J]. 地球化学, 25(6): 536-544.
张干, 盛国英, 彭平安, 郑洪汉, 邹世春. 2000. 南极乔治王岛菲尔德斯半岛湖相沉积物的分子有机地球化学特征[J]. 科学通报, 45: 2758-2762.
References:
ABRANTES F., LEBREIRO S., RODRIGUES T., GIL I., BARTELS-JÓNSDÓTTIR H., OLIVEIRA P., KISSEL C., GRIMALT J O. 2005. Shallow-marine sediment cores record climate variability and earthquake activity off Lisbon (Portugal) for the last 2000 years[J]. Quaternary Science Reviews, 24(23-24): 2477-2494.
BAC M G., BUCK K R., CHAVEZ F P., BRASSELL S C. 2003. Seasonal variation in alkenones, bulk suspended POM, plankton and temperature in Monterey Bay, California: Implications for carbon cycling and climate assessment[J]. Organic Geochemistry, 34: 837-855.
BARD E., ROSTEK F., SONZOGNI C. 1997. Interhemispheric synchrony of the last deglaciation inferred from alkenone palaeothermometry[J]. Nature, 385: 707-710.
BEADLE L. 1959. Osmotic regulation in the relation to the classification of brackish and inland saline waters[J]. Archivo Di Oceanografia E Limnologia, XI: 143-151.
BENTALEB I., FONTUGNE M., BEAUFORT L. 2002. Long-chain alkenones and U variability along a south-north transect in the Western Pacific Ocean[J]. Global Planet Change, 34: 173-183.
BRASSELL S C., EGLINTON G., MARLOWE I T., PFLAUMANN U., SARNTHEIN M. 1986. Molecular stratigraphy: a new tool for climatic assessment[J]. Nature, 320: 129-133.
CACHO I., PELEJERO C., GRIMALT J O., CALAFAT A., CANALS M. 1999. C37 alkenone measurements of sea surface temperature in the Gulf of Lions (NW Mediterranean)[J]. Organic Geochemistry. 30: 557-566.
CHU G Q., SUN Q., LI S Q., ZHENG M P., JIA X X., LU C F., LIU J Q., LIU T S. 2005. Long-chain alkenone distributions and temperature dependence in lacustrine surface sediments from China[J]. Geochimica et Cosmochimica Acta, 69(21): 4985-5003.
CONTE M H., THOMPSON A., EGLINTON G., GREEN J C. 1995. Lipid biomarker diversity in the coccolithophorid Emiliania huxleyi (prymnesiophyceae) and the related species Gephyrocapsa oceanica[J]. Phycology, 31: 272-282.
CONTE M H., THOMPSON A., LESLEY D., HARRIS R P. 1998. Enetic and physiological influences on the alkenone/alkenoate versus growth temperature relationship in Emiliania huxleyi and Gephyrocapsa oceanica[J]. Geochimica et Cosmochimica Acta, 62: 51-68.
CONTE M H., VOLKMAN J K., EGLINTON G. 1994. Lipid biomarkers of the Haptophyta[M]. The Haptophyte Algae (eds. J. C. Green and B. S. C. Leadbeater). Oxford University Press, 351-377.
CONTE M H., WEBER J C., KING L L., WAKEHAM S G. 2001. The alkenone temperature signal in western North Atlantic surface waters[J]. Geochimica et Cosmochimica Acta, 65: 4275-4287.
COOLEN M J L., MUYZER G., RIJPSTRA W I C., SCHOUTEN S., VOLKMAN J K., DAMSTÉJ S S. 2004. Combined DNA and lipid analyses of sediments reveal changes in Holocene haptophyte and diatom populations in an Antarctic lake[J]. Earth and Planetary Science Letters, 223(1-2): 225-239.
CRANWELL P A. 1985. Long-chain unsaturated ketones in recent lacustrine sediments[J]. Geochimica et Cosmochimica Acta, 49: 1545-1551.
D'ANDREA W J., HUANG Y. 2005. Long chain alkenones in Greenland lake sediments: Low δ13C values and exceptional abundance[J]. Organic Geochemistry, 36(9): 1234-1241.
DUFF K E., ZEEB B A., SMOL J P. 1995. Atlas of Chrysophycean Cysts[M]. Kluwer Academic Publishers, Dordrecht, the Netherlands.
FREEMAN K H., WAKEHAM S G. 1992. Variations in the distributions and isotopic composition of alkenones in Black Sea particles and sediments[J]. Organic Geochemistry, 19: 277-285.
FU Ming-yi, LIU Wei-guo, LI Zhong-xiang, XU Li-ming, WANG Zheng, AN Zhi-sheng. 2008. The distribution of long-chain alkenones in modern lacustrine sediments in the Lake Qinghai and lakes from the Qaidam Basi[J]. Journal of Lake Science, 20(3): 285-290(in Chinese with English abstract).
HAMMER U T. 1986. Saline lake ecosystems of the world[M]. The Netherlands: Kluwer Academic Publishers Group.
HARADA N., SHIN K. H., MURATA A., UCHIDA M., TOMOKO N. 2003. Characteristics of alkenones synthesized by a bloom of Emiliania huxleyi in the Bering Sea[J]. Geochim. Cosmochim. Acta, 67: 1507– 1519.
INNES H E., BISHOP A N., FOX P A., HEAD I M., FARRIMOND P. 1998. Early diagenesis of bacteriohopanoids in recent sediments of Lake Pollen, Norway[J]. Organic Geochemistry, 29: 1285-1295.
JARAULA C M B., BRASSELL S C., MORGAN-KISS R M., DORAN P T., KENIG F. 2010. Origin and tentative identification of tri to pentaunsaturated ketones in sediments from Lake Fryxell, East Antarctica[J]. Organic Geochemistry, 41(4): 386-397.
KIENAST M., STEINKE S., STATTEGGER K., CALVERT S E. 2001. Synchronous tropical South China Sea SST change andGreenland warming during deglaciation[J]. Science, 291: 2132-2134.
LI J G., PHILP R P., PU F., ALLEN J. 1996. Long-chain alkenones in Qinghai lake sediments[J]. Geochimica et Cosmochimica Acta, 60: 235-241.
LIU Chuan-lian, ZHAO Quan-hong, WANG Pin-xian. 2002. Water Chemistry of the Paleogene Dongying Lake: Evidence from Fossil Assemblages and Shell Isotopes[J]. Acta Geoscientica Sinica, 23: 237-242 (in Chinese with English abstract).
LIU W G., LIU Z H., FU M Y., AN Z S. 2008. Distribution of the C37 tetra-unsaturated alkenone in Lake Qinghai China: A potential lake salinity indicator[J]. Geochimica et Cosmochimica Acta, 72(3): 988-997.
LIU Z H., HENDERSON A C G., HUANG Y S. 2006. Alkenone-based reconstruction of late-Holocene surface temperature and salinity changes in Lake Qinghai, China[J]. Geophysical Research Letters. 33: L09707. doi: 10.1029/2006GL026151.
GONG Qing-jie, WU Liang-ji, WU Shi-guo, LUO You-lang, ZHANG Gan. 1999. Detection of long-chain alkenone compounds and application of U37k values in South China Sea[J]. Geochimica, 28 (1): 51- 57 (in Chinese with English abstract). LOPEZ J F., DE OTEYZA T G., TEIXIDOR P., GRIMALT J O. 2005. Long chain alkenones in hypersaline and marine coastal microbial mats. Organic Geochemistry, 36(6): 861-872.
MARLOWE I T., BRASSELL S C., EGLINTON G., GREEN J C. 1984b. Long chain unsaturated ketones and esters in living algae and marine sediments[J]. Organic Geochemistry, 6: 135-141.
MARLOWE I T., BRASSELL S C., EGLINTON G., GREEN J C. 1990. Long-chain alkenones and alkyl alkenoates and the fossil coccolith record of marine sediments[J]. Chemical Geology, 88: 349-375.
MARLOWE I T., GREEN J C., NEAL A C., BRASSELL C., EGLINTON G., COURSE P A. 1984a. Long chain (n-C37-C39) alkenones in the Prymnesiophyceae Distribution of alkenones and other lipids and their taxonomic significance[J]. British Phycological Journal, 19: 203-216.
MARLOWE I T., BRASSELL S C., EGLINTON G., GREEN J C. 1984b. Long chain unsaturated ketones and esters in living algae and marine sediments[J]. Organic Geochemistry, 6: 135-141.
MERCER J L., ZHAO M X., COLMAN S M. 2004. Seasonal variations of alkenones and UK37 in the Chesapeake Bay water column[J]. Estuarine, Coastal and Shelf Science, 63(4): 675-682
MÜLLER P J., KIRST G., RUHLAND G., STORCH I V., ROSELL-MELÉ A. 1998. Calibration of the alkenone palaeotemperature index U based on core-tops from the eastern South Atlantic and the global ocean(60°C -60°C)[J]. Geochimica et Cosmochimica Acta, 62: 1757-1772.
PEARSON E J., JUGGINS S., FARRIMOND P. 2008. Distribution and significance of long-chain alkenones as salinity and temperature indicators in Spanish saline lake sediments[J]. Geochimica et Cosmochimica Acta, 72(16): 4035-4046.
PELEJERO C., GRIMALT J O. 1997. The correlation between the U index and sea surface temperatures in the warm boundary: The South China Sea[J]. Geochimica et Cosmochimca Acta, 61: 4789-4797.
PRAHL F G., PILSKALN C H., SPARROW M A. 2001. Seasonal record for alkenones in sedimentary particles from the Gulf of Maine[J]. Deep Sea Research: part I, 48: 515-528.
PRAHL F G., WAKEHAM S G. 1987. Calibration of unsaturation patterns in long-chain ketone compositions for paleotemperature assessment[J]. Nature, 330: 367-369.
PRAHL F G., MUEHLHAUSEN L A., ZAHNLE D L. 1988. Further evaluation of long-chain alkenones as indicators of paleoceanographic conditions[J]. Geochimica et Cosmochimica Acta, 52: 2303–2310.
PRAHL F G., COWIE G L., DE LANGE G J., SPARROW M A. 2003. Selective organic matter preservation in 'burn-down' turbidites on the Madeira abyssal plain[J]. Paleoceanography, 18(2): 1052.
RONTANI J F., BEKER B.,VOLKMAN J. 2004. Long-chain alkenones and related compounds in the benthic haptophyte Chrysotila lamellosa Anand HAP17[J]. Phytochemistry, 65: 117-126.
ROSELL-MELE’A. 1998. Interhemispheric appraisal of the value of alkenone indices as temperature and salinity proxies in highlatitude locations[J]. Paleoceanography, 13: 694-703.
ROSELL-MELE’A., JANSEN E., WEINELT M. 2002. Appraisal of a molecular approach to infer variations in surface ocean freshwater inputs into the North Atlantic during the last glacial[J]. Global and Planetary Change, 34: 143-152.
ROSELL-MELÉ A., COMES P., MÜLLER P., ZIVERI P. 2000. Alkenone fluxes an anomalous UK37′ values during 1989-1990 in the Northeast Atlantic (48°N 21°W)[J]. Marine Chemistry, 71: 251-264.
ROSELL-MELÉ A., EGLINTON G., PFLAUMANN U., SARNTHEIN M. 1995. Atlantic core-top calibration of the UK37′ index as a sea-surface paleotemperature indictor[J]. Geochimica et Cosmochimica Acta, 59: 3099-3107.
ROSTEK F., RUHLAND G., BASSINOT F C., MULLER P J., LABEYRIE L D., LANCELOT Y., BARD E. 1993. Resconstructing sea surface temperature and salinity using18O and alkenone records[J]. Nature, 364: 319-321.
SACHS J P., LEHMAN S J. 1999. Subtropical North Atlantic temperatures 60,000 to 30,000 years ago[J]. Science, 286: 756-759.
SAWADA K., HANDA N., SHIRAIWA Y., DANBARA A., MONTANI S. 1996. Long-chain alkenones and alkyl alkenoates in the coastal and pelagic sediments of the northwest North Pacific with special reference to the reconstruction of Emiliania huxleyi and Gephyrocapsa oceanica ratios[J]. Organic Geochemistry, 24: 751-764.
SCHULZ H M., SCHÖNER A., EMEIS K C. 2000. Long-chain alkenone patterns in the Baltic Sea—An ocean-freshwater transition[J]. Geochimica et Cosmochimca Acta, 64: 469–477.
SEKI O., KAWAMURA K., IKEHARA M., NAKATSUKA T., OBA T. 2004. Variation of alkenone sea surface temperature in the Sea of Okhotsk over the last 85 kyrs[J]. Organic Geochemistry, 35(3): 347-354.
SHENG G Y., CAI K Q., YANG X X., LU J L., JIA G D., PENG P A., FU J M. 1998. Long-chain alkenones in Hotong Qagan Nur Lake sediments and its paleoclimatic implications[J]. Chinese Science Bulletin, 44(3): 259-263.
SHI Xing-he, ZHAO Yan-ning, DAI Sheng, XU Liang, LI Ying-ye, JIA Hong-li, ZHANG Qing-mei. 2005. Research on climatic change of Qaidam Basin since 1961[J]. Journal of Desert Research, 25(1): 123-128 (in Chinese with English abstract).
SIKES E L., FARRINGTON J W., KEGWIN L D. 1991. Use of alkenone unsaturation ratio (UK37′) to determine past sea surface temperatures: Core-top SST calibrations and methodology considerations[J]. Earth and Planetary Science Letters, 104: 36-47.
SIKES E L., LEARY T O., NODDER S D., VOLKMAN J K. 2005. Alkenone temperature records and biomarker flux at the subtropical front on the chatham rise, SW Pacific Ocean[J]. Deep Sea Research, Part I: Oceanographic Research Papers, 52(5): 721-748.
SIKES E L., VOLKMAN J K. 1993. Calibration of alkenone unsaturation ratios (Uk'37) for paleotemperature estimation in cold polar waters[J]. Geochimica et Cosmochimica Acta, 57(8): 1883-1889.
SIKES E. L, VOLKMAN J K., ROBERTSON L G., PICHON J J. 1997. Alkenones and alkenes in surface water and sediments of the Southern Ocean: Implications for paleotemperature estimation in polar regions[J]. Geochimica et Cosmochimica Acta, 61: 1495–1505.
SIKES E L., SICRE M A. 2002. Relationship of the tetra-unsaturated C37alkenone to salinity and temperature: Implications for paleoproxy applications[J]. Geochemistry Geophysics Geosystems, 3: 1063.
SONZOGNI C., BARD E., ROSTEK F., LAFONT R., ROSELL-MELE A., EGLINTON G. 1997. Core-top calibration of the alkenone index vs sea surface temperature in the Indian Ocean[J]. Deep Sea Research Part II: Topical Studies in Oceanography, 44: 1445-1460.
SUN Qing, CHU Guo-qiang. 2002. Progress in the study of temperature proxy of alkenone unsaturation[J]. Geology Geochemistry, 30(4): 63-67 (in Chinese with English abstract).
SUN Q., CHU G Q., LIU G X., LI S., WANG X H. 2007. Calibration of alkenone unsaturation index with growth temperature for a lacustrine species, Chrysotila lamellose (Haptophyceae)[J]. Organic Geochemistry, 38(8): 1226-1234.
SUN Q., CHU G Q., LI S Q., LV C F., ZHENG M P. 2004. Long-chain alkenones in sulfate lakes and its paleoclimatic implications[J]. Chinese Science Bulletin, 49(19): 2082-2086. SUN Zhen-cheng, YANG Ge-lian, QIAO Zi-zhen, YANG Fan, LI Dong-ming, JING Min-chang. 2002. Characteristics and geologic significance of calcareous nanofossils in sediments of terrestrlal salinized lakes[J]. Journal of Palaeogeography, 4: 56-63 (in Chinese with English abstract).
TERNOIS Y., SICRE M A., BOIREAU A., CONTE M H., E G. 1997. Evaluation of long-chain alkenones as paleo-temperature indicators in the Mediterranean Sea[J]. Deep Sea Research, 44: 271-286.
THEISSEN K M., ZINNIKER D A., MOLDOWAN J M., DUNBAR R B., ROWE H D. 2005. Pronounced occurrence of long-chain alkenones and dinosterol in a 25,000-year lipid molecular fossil record from Lake Titicaca, South America[J]. Geochimica Cosmochimica Acta, 69: 623-636.
THIEL V., JENISCH A., LANDMANN G., REIMER A., MICHAELIS W. 1997. Unusual distributions of long-chain alkenones and tetrahymanol from the highly alkaline Lake Van, Turkey[J]. Geochimica Cosmochimica Acta, 61: 2053-2064.
TONEY J L., HUANG Y S., FRITZ S C., BAKER P A., GRIMM E., NYREN P. 2010. Climatic and environmental controls on the occurrence and distributions of long chain alkenones in lakes of the interior United States[J]. Geochimica et Cosmochimica Acta, 74(5): 1563-1578.
VERSTEEGH G J M., RIEGMAN R., DE LEEUW J W., (FRED) JANSEN J H F. 2001. Uk37′ value for Isochrysis galbana as a function of culture temperature, light intensity and nutrient concentrations[J]. Organic Geochemistry, 32: 785-794.
VOLKMAN J K., BARRETT S M., BLACKBURN S I., SIKES E L. 1995. Alkenone in Gephyrocapsa oceanica: implications for studies of paleoclimate[J]. Geochimica Cosmochimica Acta, 59: 513-520.
VOLKMAN J K., EGLINTON G., CORNER E D S. 1980. Novel unsaturated straight-chain C37-C39 methyl and ethyl ketones in marine sediments and a coccolithophorid Emiliania huxleyi[M]. Advances in Organic Geochemistry (eds. A. G. Doulkas and J. R. Maxwell), Pergamon, New York, 219-227.
VOLKMAN J K , EGLINTON G., CORNER E D S., SARGENT J R. 1980. Novel unsaturated straight-chain C37-C39 methyl and ethyl ketones in marine sediments and a coccolithophore Emiliania huxleyi[J]. Physics and Chemistry of the Earth 12: 219-227.
WANG R L., ZHENG M P. 1998. Occurrence and environmental significance of long-chain alkenones in Tibetan Zabuye salt lake, S W China[J]. Int. Salt Lake Research, 6: 281-302.
WANG X H. 2009. Use of the Alkenone Unsaturation Ratio U37k’to Determine Past Temperature for Sihailongwan Maar(Northeast China) During the Last 1500 Years[D]. Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing.
XU Yu-lin, SUN Zhen-cheng. 1998. Discovery of the quaternary calcareous nannofossils in the deposits of interior saline lakes from northwestern China and its paleoenvironmental significance[J]. Journal of Graduate School. China University of Geosciences, 12: 49-55 (in Chinese with English abstract).
YANG Xue-xian, SHENG Guo-ying, LU Jia-can, FU Jia-mo. 1996. Characteristics and paleoenvironmental significance of biomarks in sediments from Hotong Qagan Num (soda lake)[J]. Geochimica, 25(6): 536-544(in Chinese with English abstract).
YAMAMOTO M., SHIRAIWA Y., INOUYE I. 2000. Physiological responses of lipids in Emiliania huxleyi and Gephyrocapsa oceanica (Haptophyceae) to growth status and their implications for alkenone paleothermometry[J]. Organic Geochemistry, 31(9): 799-811.
ZHANG G., SHENG G Y., PENG P A., ZHENG H H. 2000. Molecular organic geochemical peculiarities of lacustrine core sediments in Fildes Peninsula, King George Island, Antarctica[J]. Chinese science bulletin, 45: 67-70.
ZHAO M X, EGLINTON G., READ G. 2000. An alkenone (U) quasi-annual sea surface temperature record (A.D.1440 to 1940) using varved sediments from the Santa Barbara Basin[J]. Organic Geochemistry, 31: 903-917.
ZINK K G., LEYTHAEUSER D., MELKONIAN M., SCHWARK L. 2001. Temperature dependency of long-chain alkenone distributions in recent to fossil limnic sediments and in lake waters[J]. Geochimica et Cosmochimica Acta, 65: 253-265.
The Occurrence and Distribution of Long Chain Alkenones in Lakes
SUN Qing1), CHU Guo-qiang2), LIU Guo-xiang3), WANG Xiao-hua2), LIU Mei-mei1), SHI Li-ming1), XIE Man-man2), LIN Yuan1)
1) National Research Center of Geoanalysis, Beijing 100037;
2) Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing 100029;
3) Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, Hubei 430072
The alkenone unsaturation paleothermometer has been successfully and widely applied to reconstructing SST in most oceanographic settings. The utility of the alkenone unsaturation index in the marine system has stirred the interest in the study of LCK in the limnic system, in the hope that it can also be used as a paleotemperature proxy in lacustrine sediments. A suite of long-chain di-, tri- and tetra-unsaturated ketones whose chain lengths range from C37 to C39 with variable patterns of LCK were detected in the lake sediment samples. The ratio of C37:4methyl ketone to the sum of C37 alkenones observed in different lakes is highly variable, and higher than that seen in marine systems. Different distribution patterns of LCK were detected in the surface sediment samples. The very high ratio of C37 / C38 may imply that dominant LCK-producing algae are probably similar to the species C. lamellosa and I. galbana. The detection of C38:4-2 ethyl alkenones suggests that the precursor organism might be similar to E. huxleyi G. oceanica and some species of Isochrysis galbana. However, if only C38:4-2 ethyl alkenonewas identified, the precursor organism may be or closely related species to Chrysotila lamellosa or some species of Isochrysis galbana. Empirical relationships between the alkenone unsaturation index U37k’and different temperature sets (mean annual air temperature, mean annual air temperature in different seasons, and lake surface water temperature of July) were tested. The better correlation between U37k’and temperature was obtained using mean annual air temperature, mean air temperatures of spring and autumn. Based on the new data from the reference the authors fit a new global linear regression of U37k’and MAAT can be expressed as U37k’= 0.031× T + 0.094 (n=76, r2= 0.67), it covers the lake system from the lakes in Greenland to the lakes in the northern part of tropical area in China. Although problems such as species-uncertainty and other unknown factors for U37k’temperature dependence remain existent, the equation might be the representative of the average contribution of LCK to sediments for these data over a wide range of surface temperatures, water chemistry and different alkenones-producer algal populations. A lacustrine source, the non-calcifying species Chrysotila lamellosa Anand (Haptophyceae), was collected and isolated from an inland saline water body, Lake Xiarinur (Inner Mongolia). Its alkenone distribution pattern is similar to that of coastal marine strains of C. lamellosa, but the relationship between U37k’index and culture temperature for the lacustrine species is quite different from that of the coastal species. A significant feature of the alkenones in this strain of C. lamellosa is the lack of C38 methyl alkenones, which might be used to distinguish the species from the marine haptophyte species Emiliania huxleyi and Gephyrocapsa oceanica. The authors examined U37k’and U37kvalues for C. lamellosa as a function of culture temperature in a batch culture experiment. The calibration for U37k’versus culture temperature (T) was U37k’= 0.0257× T -0.2608 (n = 9, r2= 0.97), from 14°C to 22°C. The slope of the equation is similar to the empirical relationship between U37k’and mean annual air temperature in saline lakes. The authors’ studies show that the alkenone unsaturation index U37k’is strongly controlled by environmental temperature the precursor organisms live in, and can be used for palaeoclimate reconstruction. This supports the suggestion that the biosynthetic pathway of alkenones and the mechanism of their temperature signal may be similar in both marine and limnic systems. LCK might be used as an important paleotemperature proxy in the limnic environment.
lake; alkenone; U37k’; distribution pattern; temperature; precursor organism
K928.43; O622
A
1006-3021(2010)04-485-10
本文由国家自然科学基金项目(编号: 40572101; 40972121; 40102016)和国土资源部百人计划项目联合资助。获中国地质科学院2009年度十大科技进展第四名。
2010-06-10; 改回日期: 2010-07-08。
孙青, 女,1967年生。研究员。主要从事古气候变化研究。通讯地址: 100037, 北京市西城区百万庄大街26号。电话: 010-68999590。E-mail:sunqingemail@yahoo.com.cn。
