香料烟青枯病抗性基因的遗传分析
2010-07-31高加明王志德张兴伟刘艳华牟建民
高加明,王志德,张兴伟,刘艳华,牟建民,任 民,徐 军
(1.农业部烟草类作物质量控制重点实验室,中国农业科学院烟草研究所,青岛 266101;2.中国农业科学院研究生院,北京100081)
青枯病是一种由假单胞杆菌属细菌(Ralstonia solanacearum)引起的一种破坏性极强的土传病害,是典型的维管束病害,最显著的症状是枯萎。烟草青枯病是烟草的重要病害,一旦发病即可造成全株死亡,对烟叶生产造成极大的经济损失。目前,烟草生产上主要采取药剂、生物以及轮作等方式防治青枯病,但各种防治措施都存在着各自的不足及局限性,因而无法从根本上进行防治。因此,明确烟草青枯病抗性的遗传规律,以便采取正确的育种手段,培育抗病新品种是一种最有效的防治措施[1]。
近年来有关青枯病研究开展最多的是茄科植物,如番茄、茄子[2]、马铃薯[3]等。研究表明,青枯病的抗性遗传规律有隐性遗传、加性效应为主的不完全显性等,甚至有研究认为抗性在幼龄植株中表现不完全显性,在成株期则为隐性等不同观点。关于基因的个数也无定论,大多数认为受多基因控制,但也有不少研究者认为是受显性单基因控制[4]。
烟草青枯病有几种不同水平的抗源,大部分是中抗以下品种,很少有高抗青枯病的品系,在野生烟属种中也未鉴定出抗病性[5]。所以从现有抗源中研究遗传机理,准确了解烟草青枯病抗性基因的遗传机理,为培育出高抗病性的烟草品种提供一定的理论依据。作者采用章元明[6]植物数量性状混合遗传模型主基因+多基因多世代联合分析方法,研究了香料烟Xanthi×Samsun组合的F2分离群体的病情指数的遗传规律,为优质抗病香料烟新品种的选育提供依据[7]。
1 材料和方法
1.1 供试材料
选用青枯病抗病品种 Xanthi和感病品种Samsun配置杂交组合。2008年在中国农业科学院烟草研究所即墨基地种植亲本,进行杂交,收获F1种子。并于当年9月份在温室加代,套袋自交,翌年2月收获F2种子。
1.2 试验设计
将亲本、F1和 F2种于福建省三明市青枯病病圃,此病圃是国家定点青枯病抗性鉴定病圃,调查烟草青枯病病情。试验设3个重复,完全随机区组设计,3个小区,每个小区40株。对照品系栽在中间,设抗病品种 D101,感病品种长脖黄为对照。按当地优质烟叶生产技术规范进行田间管理。
1.3 青枯病病情调查
由当地试验技术人员按YC/T39-1996行业标准进行调查[8],分别调查3次,以发病高峰期的调查数据作为分析数据,并计算病情指数。
病情指数=∑(各级病株数×该病级值)/(调查总株数×最高级值)×100
1.4 数据分析
采用章元明[6]植物数量性状混合遗传模型主基因+多基因多世代联合分析方法对 Xanthi×Samsun组合P1、P2、F1和F2等4个世代单株的病情指数进行分析,通过比较1对主基因(A),2对主基因(B),多基因(C),1对主基因+多基因(D)和 2对主基因+多基因(E)等5类24种遗传模型的AIC值,并进行遗传模型的适合性测验,包括均匀性检验(U12,U22和U32),Smirnov检验(nW2)和Kolmogorov 检验(Dn),综合考虑极大似然函数、AIC值和适合性检验的结果确定最优模型。最后估算最适合模型的主基因和多基因效应值、遗传率等一阶遗传参数和二阶遗传参数等[9]。
在主基因+多基因的混合遗传模型中,表现型值(p)表示为群体平均数(m)、主基因效应(g)、多基因效应(c)和环境效应(e)之和,即p=m+g+c+e,因此,表现型方差(σp2)可表示为主基因方差(σmg2)、环境方差(σe2)和多基因方差(σpg2)之和,即σp2=σ e2+σmg2+σpg2。分离群体的主基因方差(σmg2)等于表现型方差(σp2)减去纯合主基因型成分分布方差。多基因方差(σpg2)为纯合主基因型成分分布方差减去环境方差(σe2)。主基因遗传率 hmg2(%)=σmg2/σp2×100%;多基因遗传率 hpg2(%)=σpg2/σp2×100%。
2 结 果
2.1 4世代田间病情调查结果
对亲本、F1和F2的调查结果见表1。结果表明,3个重复之间的病情差异不显著。P1是感病品种,病情指数为 77.50;P2是抗病品种,病情指数为13.75。杂交获得的 F1代为中抗型,其病情指数略高于亲本的平均值。后代F2的表型分布呈现双峰偏态分布,作者认为它有可能既有主效基因的作用,又有微效基因的作用。
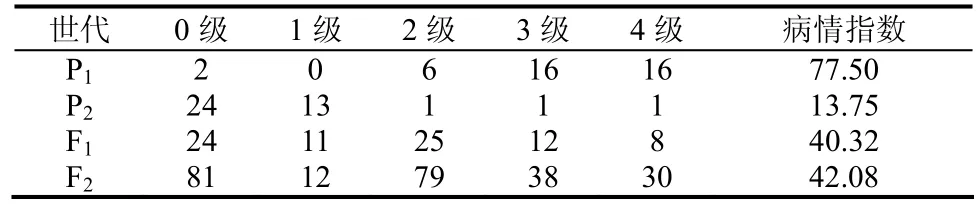
表1 4世代烟草青枯病的病情分布Table 1 Disease distribution for four generations of tobacco bacterial wilt
2.2 田间抗性的统计遗传分析
用4世代联合分离分析方法,通过IECM算法获得1对主基因(A),2对主基因(B),多基因(C),1对主基因+多基因(D)和 2对主基因+多基因(E)等 5类24种遗传模型的极大似然函数值和AIC值,见表2。根据AIC值准则,AIC值小的遗传模型为可能最佳模型。从而选出AIC值小的E-1、E-3、E-4、E-5和E-6作为备选模型。然后对其5个备选模型进行适合性(U12、U22、U32、nW2和Dn)检验,结果见表3。选择统计量达到显著水平个数较少的模型作为最优模型。由表3可知,20个适合性检验统计量中,E-1模型中只有2个达到显著水平(α=0.05),即有2个适合性检验统计量表明E-1模型与群体的分布是不一致的[10];其它4个模型中均有11个达到显著水平(α=0.05)。因此选择 E-1模型,即 2对加性-显性-上位主基因+加性-显性多基因混合模型为烟草青枯病抗性遗传的最优模型。
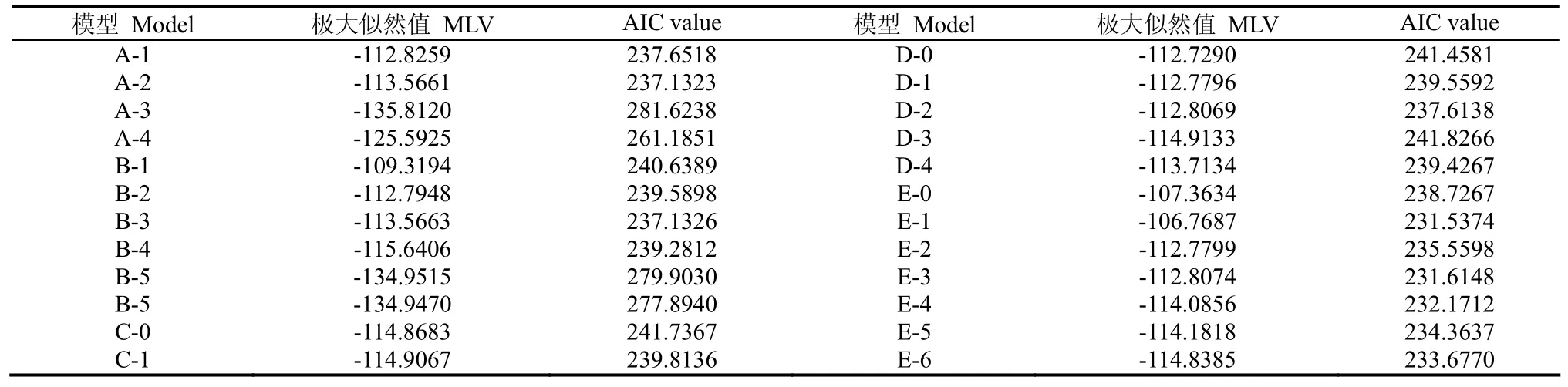
表2 Xanthi×Samsun组合青枯病抗性在不同遗传模型下的MLV和AIC值Table 2 Maxinum likelihood value(MLV) and Akaike’s information criterion(AIC) for resistance to bacterial wilt in the cross of Xanthi×Samsun under different genetic models
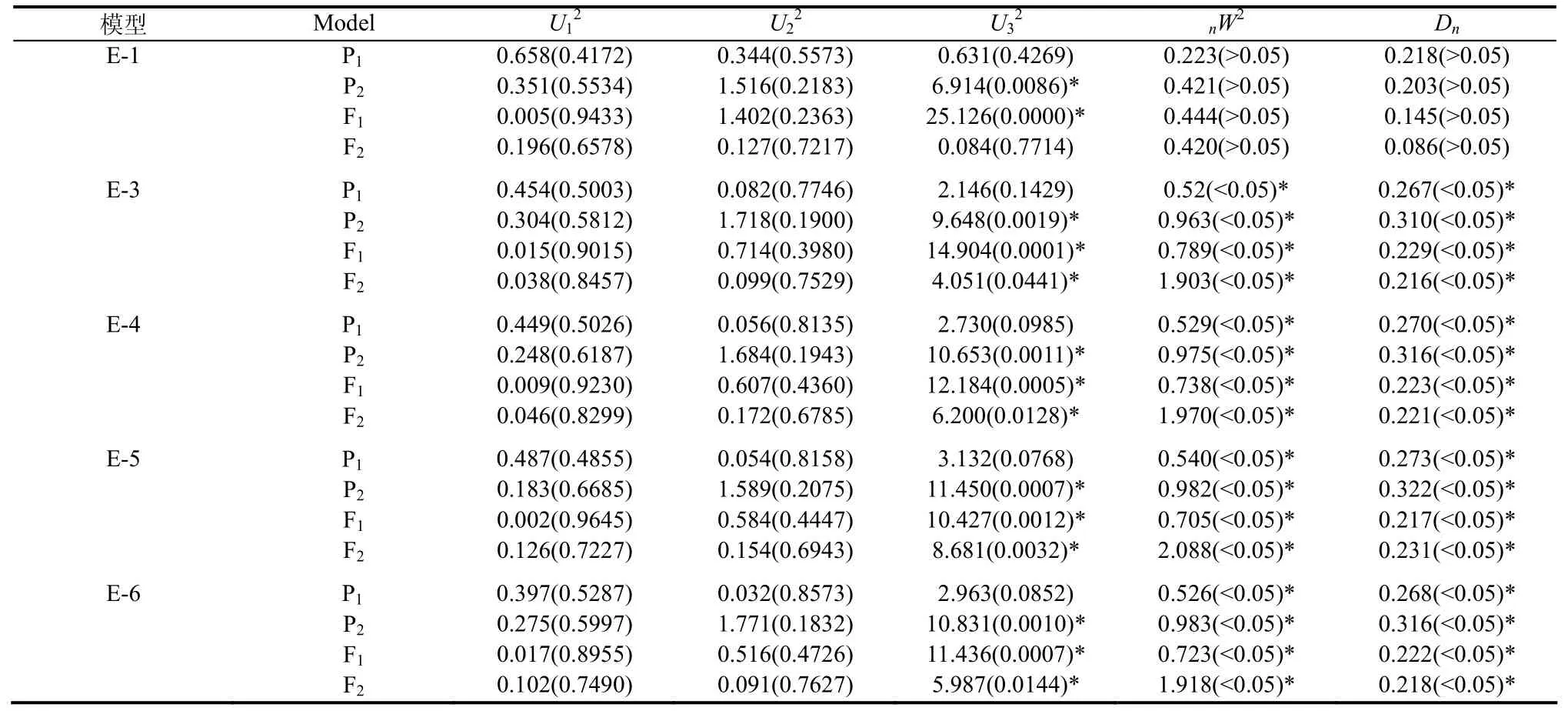
表3 Xanthi×Samsun组合青枯病抗性遗传模型的适合性检验Table 3 Tests on goodness-of-fit of genetic models for resistance to bacterial wilt in the cross of Xanthi×Samsun
2.3 遗传参数的估计
此模型遗传参数的估计见表4。从一阶遗传参数可以看出烟草青枯病抗性基因2对主基因的加性效应∣da∣≈∣db∣,说明 2对主基因的加性效应几乎相等,但是符号相反,几乎可以抵消;2对主基因的显性效应∣ha∣=∣hb∣,即显性效应相等;2对主基因显性度的绝对值∣ha/da∣、∣hb/db∣都小于1,说明2对主基因的加性效应大于显性效应,2对主基因都以加性效应为主;2对主基因间的加性互作为负值,说明两基因同时存在时植株容易感病;第1对主基因的显性效应×第2对主基因的加性效应大于第1对主基因的加性效应×第2对主基因的显性效应,并且同为正值,说明育种中更应该注重第1对主基因的显性效应×第2对主基因的加性效应之间的互作,更利于提高后代的抗病能力;2对主基因的显性互作为0.2076;多基因的加性效应大于主基因的加性效应,说明多基因的影响还是比较大的。
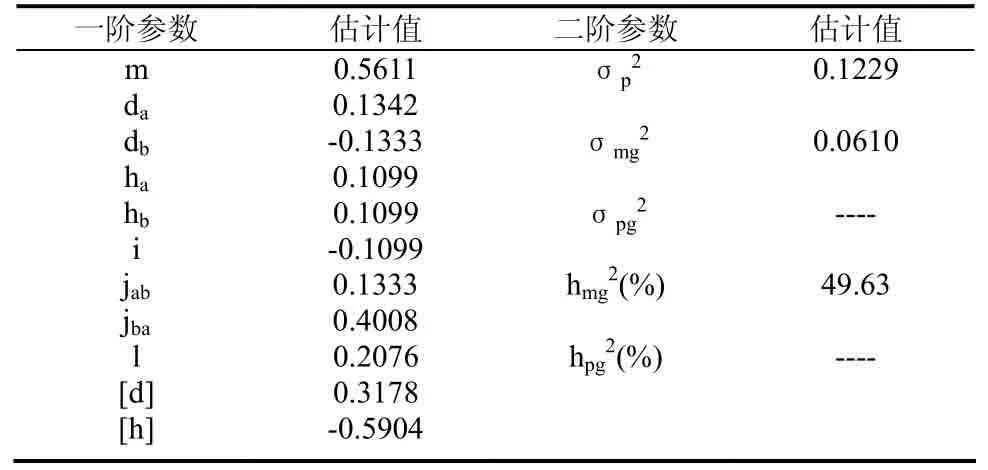
表4 最适遗传模型E-1下抗性基因的遗传参数估计Table4 The estimates of genetic parameters for resistant gene under the E-1 model
二阶遗传参数,主基因的遗传率为49.63%;由于环境误差影响很大,导致多基因方差为负值,说明烟草青枯病的发生受环境因素影响比较大,今后应该从试验设计、田间种植等方面减小环境因素的影响,估计出准确的多基因遗传率。
3 讨 论
前人研究表明,对于烟草青枯病抗性的遗传机理还没有统一的定论。Matsuda和Ohashi (1973)报道Awa、Hatano、Kokubu 与Odaruma 等中抗品种的抗性受部分显性基因Rps控制,高抗品种Enshu、Hatanodaruma的抗性不仅受Rps基因控制,还受多基因影响[11]。在国内,杨友才等[4]通过使用强致病力的单一菌株,对烟草抗性资源 Ti448A的抗性遗传机理进行了研究,结果表明烟草青枯病抗性是受显性单基因控制,并从分子水平上进行了进一步的验证,结果抗病与感病的分离比为3:1,符合 1对显性基因控制的遗传规律。出现不同结果可能是由于以下原因:一是试验所选的材料不同,不同抗源的遗传机理不同;二是病原菌的不同,青枯菌菌系分化比较复杂,当在田间自然发病时,只有这些位点上的抗性基因均为显性时,才表现为抗病,若一处为隐性,则有可能表现为感病,这样就会导致病情的差异;三是环境的不同,从分析结果也可以看出环境对烟草青枯病有很大的影响,不同环境导致病情的分化很大。
本研究通过对香料烟品种 Xanthi×Samsun组合P1、P2、F1和F2等4个世代采用主基因+多基因混合遗传模型进行了联合分析,结果表明烟草青枯病抗性是受 2对加性-显性-上位主基因+加性-显性多基因(E-1模型)控制遗传,与前人研究比较一致。Matsuda[11]认为Xanthi的抗性不仅由部分显性基因控制,还包括多基因的影响。但无论是主基因、多基因的加性、显性效应,还是主基因间的互作效应都非常小;从主基因的遗传率来看,烟草青枯病的发生受环境因素的影响比较大。
本研究对表型数据进行了分析,其结果是初步的。Nishi 等[1]利用来自白肋烟品种间杂交(W6×Michinoku)所产生的125 个株系的DH 群体,构建了第一张白肋烟 AFLP分子遗传连锁图。将W6中抗青枯病的一个QTL定位到一个长32cM含有 15个标记的连锁图谱上。此位点可解释的表型变异高于30%。随着SSR引物的不断开发,目前正在利用SSR、SRAP等分子标记来构建遗传连锁图谱,从而对青枯病抗性进行基因定位,进而获得与目标基因紧密连锁的分子标记,为进行分子标记辅助选择(Marker-Assisted Selection,MAS)培育新品种提供了可能[12]。
[1]Nishi T,Tajima T,Noguchi S,et al.Identification of DNA markers of tobacco linked to bacterial wilt resistance[J].Theor Appl Genet,2003(106):765-770.
[2]朱华武,姚元干,刘志敏,等.茄子抗青枯病的遗传规律研究[J].湖南农业大学学报(自然科学版),2004,30(3):288-289.
[3]郜刚,屈冬玉,连勇,等.二倍体马铃薯群体青枯病抗性鉴定及遗传分析[J].马铃薯杂志,1998,12(4):212-216.
[4]杨友才,周清明,朱列书.烟草青枯病抗性基因的遗传分析及RAPD标记[J].中国烟草学报,2006,12(2):38-42.
[5]佟道儒.烟草育种学[M].北京:中国农业出版社,1997:446-453.
[6]章元明,盖钧镒,张孟臣.利用P1F1P2和F2或F2∶3世代联合的数量性状分离分析[J].西南农业大学学报,2000,22(1):6-9.
[7]蔡长春,张俊杰,黄文昌,等.利用DH群体分析白肋烟黑胫病抗性的遗传规律[J].烟草科技,2009(1):54-59,63.
[8]王志德,王元英,牟建民,等.烟草种质资源描述规范和数据标准[M].北京:中国农业出版社,2006:70-71.
[9]盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:中国科学出版社,2003.
[10]郑义,陆维忠,陈建民,等.大麦赤霉病新抗源盐96157抗性的遗传分析[J].麦类作物学报,2008,28(2):329-333.
[11]Mastsuda T,Ohashi Y.Inheritance of resistance to bacterial wilt resistant varieties in tobacco [J].Jap Tour Breeding,1973,23:175-180.
[12]Johnson E S,Wolff M F,Wernsman E A.Marker-Assisted Selection for Resistance to Black Shank Disease in Tobacco [J].Plant Disease,2002,86(12):1303-1309.
