不同氮磷组合对短穗兔儿草克隆繁殖特性的影响
2010-06-04周华坤杨元武赵新全
杨 帆,周华坤,叶 鑫,杨元武,赵新全
(1.青海大学农牧学院草业科学系,青海西宁 810016;2.中国科学院西北高原生物研究所,青海西宁 810001)
短穗兔耳草(Lagotisbrachystachya)分布于青藏高原海拔2 600~4 600 m地区,生长在河边滩地、弃耕地和山坡撂荒地等生境中的多年生草本植物,以有性和无性方式进行繁殖,主要进行无性繁殖即克隆繁殖,在一些人为活动所形成的次生裸地上常会成为植被恢复演替过程中的先锋植物[1],是高寒地区典型的多年生匍匐茎植物,也是高寒草甸的主要伴生种,无性繁殖能力强,是草地退化演替进程中出现的主要指示种[2-5]。短穗兔耳草的克隆繁殖主要通过直立茎基部节上生出侧向生长的1条或数条匍匐茎 ,在匍匐茎的末端产生幼苗和不定根 ,成为克隆分株;匍匐茎上分布有若干对称茎生叶,有的克隆分株也可继续产生次一级克隆分株,匍匐茎断裂后也可直接成为新个体。近年来,青藏高原草地退化形势日趋严重,退化进程中均伴有短穗兔耳草和鹅绒委陵菜的大量出现[6,7]。另外,国内对短穗兔耳草的研究主要涉及其种群生长特性、基株大小及其与克隆生长特征的关系,海拔梯度和退化程度对其无性生长和繁殖特性的影响,以及其化学成分和抑菌活性的研究等[[8-11]。但结合高寒草甸退化演替阶段和不同人为干扰对其克隆繁殖特性影响的研究报道较少。对短穗兔耳草也仅限于不同海拔高度和不同生境下克隆生长特征的调查研究[1-14]。因此,开展人工控制土壤条件下对短穗兔耳草移栽试验,直接观察其克隆分株在空间的分布格局,探讨土壤水、肥变化下表现出的可塑性,并将其用于退化生态系统的恢复与重建有着良好的前景,可为生态系统的退化、恢复与重建提供理论依据。旨在通过控制土壤中氮磷肥的施入量,进行短穗兔儿草的移栽,了解其生长及克隆繁殖特性。
1 材料和方法
1.1 试验材料及来源
试验材料为短穗兔耳草,采自于青海省果洛藏族自治州境内玛沁县。2008年5月下旬于玛沁县采挖已萌发的短穗兔耳草2~3叶植株进行移植。采集地玛沁县位于江河源区,地处 E 98°48′~100°55′,N 33°43′~35°16′,草地类型为典型的高寒草甸 ,境内地势高亢,平均海拔4 000 m。属高寒半湿润性气候,除冷暖两季外没有明显的四季之分,日照充足,历年日照平均值2 500 h。冷季持续时间7~8个月,且风大雪多;暖季湿润,长4~5个月。平均气温在0℃以下,全年无绝对无霜期。年降水量420~560 mm,多集中在5~10月[6]。
1.2 试验地概况
2008年5~10月在青海省西宁市青海大学农牧学院草业科学系统试验地。试验地处 N 36°35′,E 101°36′,海拔 2 260 m,年降水量 428.3 mm,蒸发量1 363.6 mm,年均日照2 703.2 h,平均气温 3.9~5.5℃,最高气温26.9℃,最低-18.9℃。试验总面积为106 m2,土壤为栗钙土,其基本理化性质见表1。

表1 供试土壤的基本理化性质Table 1 Basic properties of tested soil
1.3 试验设计
试验采用随机区组设计,共15个小区,每个小区面积为2.4 m2,种植间距30 cm×30 cm。试验所用氮肥为尿素,磷肥为二铵,不施 K 肥,设置 A 、B、C、D、E 5个施肥组合(表2),以处理A为对照,每个处理设置3个重复。所有肥料分3次施入,定植前作为基肥施入40%,采收前期和中后期各施入30%。总灌水量为2 000 m3/hm2,分10次灌入,不考虑降水。
1.4 观测指标及数据处理
每隔15 d观测1次。测定茎生叶数、分株叶片数、分株高、匍匐茎数、分株根长和匍匐茎长度。地上部分指标均在田间测得,地下部分在秋季采收后,测定基株根长和分株根长。取各试验处理最大观测值的平均值。
利用统计数据处理软件DPS v6.55进行方差分析和显著性检验,在差异显著时对其采用新复极差法进行多重比较。

表2 施肥处理Fable 2 Fertilization treatment design kg/hm2
2 结果与分析
2.1 不同氮磷组合对短穗兔耳草基株生长的影响
不同施肥水平对短穗兔耳草的基株叶片数的影响存在显著差异,9月25日的观察值表明,随着氮磷组合肥料的施入,基株叶片数变化较明显,其中,以处理C,即氮肥施入量为90 kg/hm2、磷肥施入量为60 kg/hm2时对基株的影响最大,平均叶片数达16.25片,而随着氮肥和磷肥施入量的增加,短穗兔儿草基株叶片数显著减少(表3),说明过量的氮肥和磷肥会抑制短穗兔耳草基株叶片数的增加。处理C与处理B、D的基株根长差异并不显著,只与处理A和E显著,说明肥料的施入对短穗兔耳草地下根长的影响并不明显。但对基株而言,叶片数和根长均是在处理C的条件下达到最大值,分别为16.25片和17.20 cm(表4)。
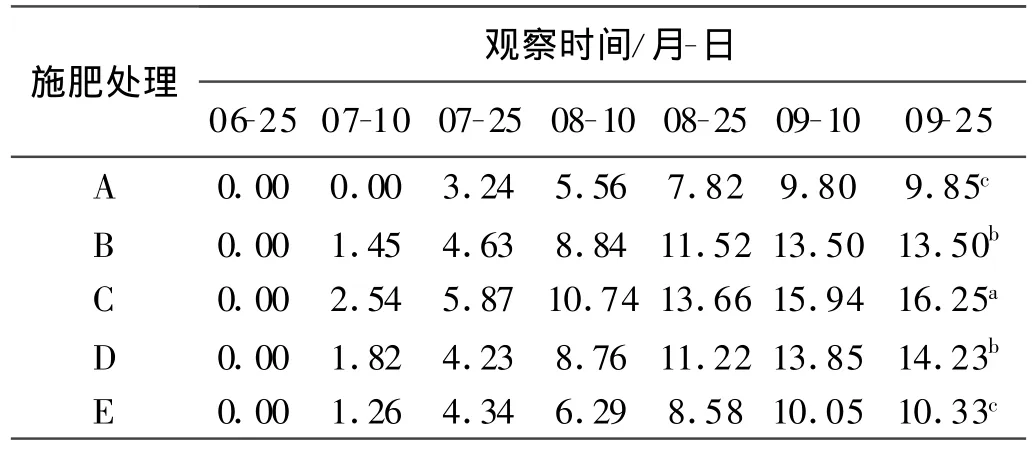
表3 不同施肥处理短穗兔耳草基株叶片数Table 3 Leaf number of genet in different treatments 片

表4 不同施肥处理短穗兔耳草基株根长Table 4 Root length of genet in different treatments cm
2.2 不同氮磷组合对短穗兔耳草克隆分株的影响
2.2.1 对短穗兔儿草匍匐茎数量的影响 不同氮磷组合在生长初期对短穗兔耳草匍匐茎数量影响并不明显(表5),在短穗兔儿草移栽后1个月开始陆续长出匍匐茎,到8月10日各处理的短穗兔耳草匍匐茎数增加较快,9月10日以后,处理A、D、E的匍匐茎数量达到最大值,往后不再增加,而处理B、C还有缓慢增加,到采收时达到最大值,9月25日的数值为各处理间匍匐茎数量差异显著,处理C的匍匐茎数最高,为7.23条。

表5 不同施肥处理短穗兔耳草匍匐茎数Table 5 Stolon number in different treatments 条
2.2.2 对短穗兔儿草匍匐茎长度和茎生叶数的影响 不同氮磷组合处理对短穗兔耳草茎生叶数的影响与对匍匐茎数的影响一致,在生长初期影响不明显,到移栽后1个月,随着匍匐茎的生长,茎生叶数逐渐增加。9月25日各处理间短穗兔儿草茎生叶片数差异显著,处理C的茎生叶片数最高为11.25片(表6)。9月25日处理C、D、E的匍匐茎长与处理A、B差异显著,但处理C、D、E间的差异并不显著,处理C的平均匍匐茎最长,达到13.36 cm(表7)。
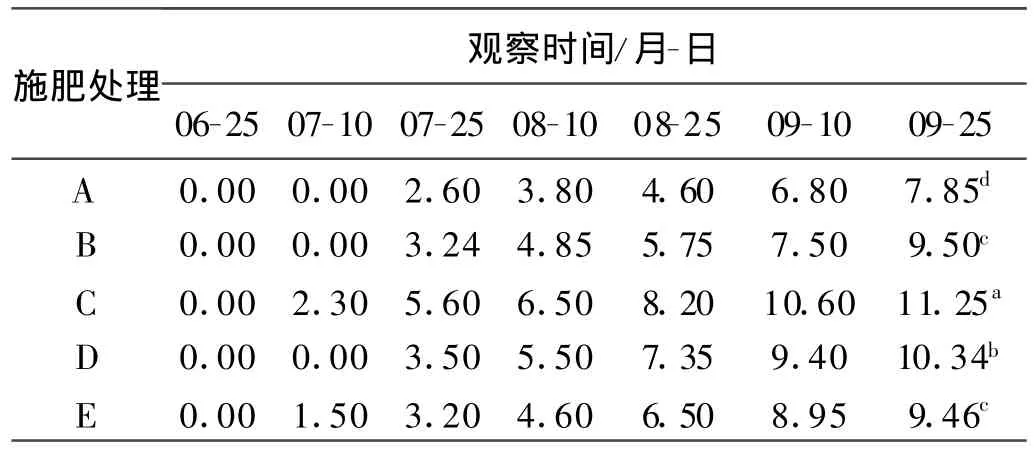
表6 不同施肥处理短穗兔耳草茎生叶数Table 6 Cauline leaf number in different treatments 片
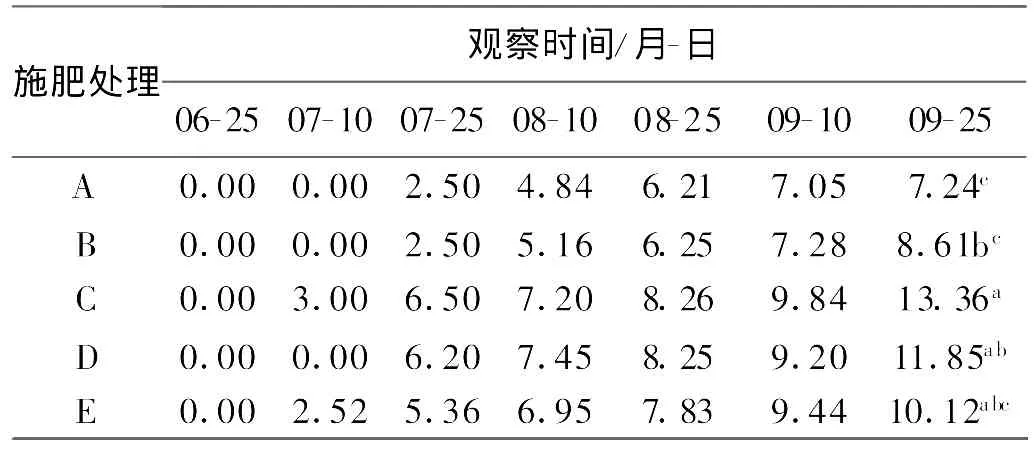
表7 不同施肥处理短穗兔耳草匍匐茎长Table 7 Stolon length in different treatments cm
2.2.3 对短穗兔儿草分株叶片数的影响 9月25日各处理差异显著,处理C和D的分株叶片数最高,随着肥料的施入量高于氮肥90 kg/hm2,磷肥60 kg/hm2时,分株叶片数开始降低(表8),说明处理C的施肥量对于增加短穗兔耳草分株叶片数有一定的作用,而高于这个水平时,分株叶片数有降低的趋势。

表8 不同施肥处理短穗兔耳草分株叶片数Table 8 Leaf number of ramet in different treatments 片
2.2.4 对短穗兔儿草分株叶片数的影响 9月25日不同氮磷组合处理间短穗兔耳草分株根长存在显著差异,以处理C为最高,达到4.32 cm,高于周华坤[6]等提出的分株根长2.73cm(表9)。这在一定程度上表明施肥可以增加短穗兔耳草克隆分株根长。

表9 不同施肥处理短穗兔耳草分株根长Table 9 Root length of ramet in different treatments cm
3 小结与讨论
试验结果表明:不同氮磷施肥组合对短穗兔耳草的克隆生长有很大的影响,当其他条件不变时,短穗兔耳草的克隆繁殖特性在不同氮磷施肥组合水平下差异均显著,且短穗兔耳草的匍匐茎数、茎生叶片数、分株叶片数和根长均在处理C时达到最大值,这些指标的变化趋势基本是随着肥力水平的提高,先增加再降低,以处理C(氮肥90 kg/hm2,磷肥60 kg/hm2)的表现最好,因此,土壤氮磷肥力条件的差异对短穗兔儿草的克隆繁殖特性存在一定的可塑性。
在贺宝珍等[12]的研究中发现,短穗兔耳草的匍匐茎数在海拔3 700 m处平均为5条,海拔3 900 m处平均为9条,在海拔4 100 m处平均为4条。而此次研究中,处理C的匍匐茎数量平均值最高为,7.23条,引起这个现象的原因还有待于进一步研究。而在周华坤等人研究中发现,在牧草生长盛期,短穗兔耳草的匍匐茎的茎生叶数为12.54片,长度为15.14 cm,略高于本研究结果。在处理C条件下,短穗兔耳草的基株叶片数为16.25片、基株根长为17.20 cm、分株叶片数可达6.12片,均高于周华坤等人提出的7.98片、9.11 cm和3.08片,这在一定程度上也说明中等施肥量可以促进短穗兔耳草的生长。但究竟是哪一种肥料起到了促进短穗兔耳草克隆繁殖生长特性,以及移栽后海拔变化对短穗兔耳草克隆繁殖特性有何影响,均有待于进行进一步的研究。在资源异质性分布的环境中,克隆植物往往通过克隆构型的可塑性变化来调整资源获取对策,具有重要的生态学意义。短穗兔耳草由于氮磷肥力梯度导致的资源差异的反应,是其对资源利用达到的最合理状态,是一种选择适应的结果。
[1]周华坤,赵亮,赵新全,等.短穗兔耳草的克隆生长特征[J].草业科学,2006(12):60-64.
[2]王刚,张大勇.生物竞争理论[M].西安:陕西科学技术出版社,1996:106-137.
[3]王艳红,王珂,邢福.匍匐茎草本植物形态可塑性、整合作用与觅食行为研究进展[J].生态学杂志,2005,24(1):70-74.
[4]刘伟,王启基,王溪,等.高寒草甸“黑土型”退化草地的成因和生态过程[J].草地学报,1999,7(4):300-307.
[5]张途正,王生耀.高寒地区微肥对多年生人工草地生产性能的影响[J].草原与草坪,2009(6)20-23.
[6]周华坤,韩发,周立,等.高寒草甸退化对短穗兔耳草克隆生长特征的影响[J].生态学杂志,2006,25(8):873-879.
[7]叶上游,潘爽,王景波,等.克隆植物生理整合作用研究进展[J].草原与草坪,2008(6):63-69.
[8]李镇清.克隆植物构型及其对资源异质性的响应[J].植物学报,1999,41(8):893-895.
[9]周华坤,周立,赵新全,等.江河源区“黑土滩”型退化草场的形成过程与综合治理[J].生态学杂志,2003,22(5):51-55.
[10]周华坤,周兴民,周立,等.鹅绒委陵菜(Potentilla anserina L.)生长特征[J].西北植物学报,2002,22(1):9-17.
[11]淮虎银,魏万红,张镱锂,等.不同海拔高度短穗兔耳草克隆生长及克隆繁殖特征[J].应用与环境生物学报,2005,11(1):18-22.
[12]贺宝珍,赵建中,周华坤.不同海拔短穗兔耳草生长特征研究[J].安徽农业科学,2008,36(26):11277-11279.
