PLptn真核载体的构建表达及其趋化活性测定
2010-06-01蒋朋飞曹金锁张德礼
柳 超,蒋朋飞,曹金锁,张德礼
(西北农林科技大学动物医学院病毒免疫与生物信息研究室,陕西杨凌712100)
趋化因子是一种能够趋化和活化白细胞的细胞因子,在炎症和免疫反应中发挥着重要的作用,现在发现的趋化因子有CXC,CC,CX3C和 C共四族。淋巴细胞趋化因子(Lptn)为趋化因子C家族成员,可由CD8+T细胞、NK细胞、肥大细胞、嗜酸性粒细胞、γ δ T 细胞等多种细胞产生,主要作用于 CD4+T细胞、CD8+T细胞、NK 细胞、B细胞、中性粒细胞等[1]。Lptn的结构、功能等在人及小鼠中研究较为深入,目前人和小鼠的Lptn单克隆抗体已实现商品化生产。Zavala-Flores L M等[2]已成功用乳酸杆菌表达了人淋巴细胞趋化因子,并在体外做了活性验证,而Lptn的研究在猪体内却未见相关报道。为此本研究构建了猪淋巴细胞趋化因子(PLptn)真核表达载体,并转染至猪脐静脉血管内皮细胞系(SUVEC),在体外测定了其趋化生物学活性,为进一步研究其功能奠定了基础。
1 材料与方法
1.1 试剂和材料 T rizol Reagent试剂、Lipofectamine TM 2 000购自Invitrogen公司,RT-PCR反转录试剂盒购自Fermentas公司,premix Taq DNA聚合酶、限制性内切酶以及T4 DNA连接酶购自 TaKaRa宝生物工程(大连)有限公司,Anti-GFP Antibody购自天根生物科技有限公司,羊抗鼠IgG/HRP购自北京博奥森生物科技有限公司,DMEM细胞培养基购自Gibco公司,6孔细胞培养板购自Costar公司,Western blot所用聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF)为美国Millipore生产。
1.2 方法
1.2.1 猪胸腺组织总 RNA提取 T rizol法从健康成年长白猪胸腺组织中提取总RNA。
1.2.2 PLptn cDNA的获得 按常规的 RT-PCR方法进行,根据GenBank登录号为NM_001134345序列设计引物,序列如下 Upper:5′-CGCAAGCTTCCATGAGACT TCTCATC-3′,Lower:5′-TAGGATCCCTACCCAGTCAGGGT T-3′。PCR 总 体系为 25 μ L,反应体系如下 :12.5 μ L Taq DNA polymerase,上下游引物(20 pmol)各 0.5 μ L,1.25 μ L反转录产物,10.25 μ L ddH2O。反应参数为:95℃5 min,30个循环(94℃30 s,56℃30 s,72℃30 s),最后72℃延伸10 min。
1.2.3 真核载体pEGFP-PLptn的构建 PCR产物纯化后与pMD-18T载体连接,构建亚克隆载体pMD-18T-PLptn,转化 DH5α感受态细胞,经氨苄抗性筛选、测序及双酶切对重组菌进行鉴定。将正确的目的基因连接至pEGFP-C1中,构建真核载体pEGFP-PLptn。
1.2.4 真核载体pEGFP-PLptn转染SUVEC 抽提真核载体pEGFP-PLptn,紫外分光光度计测定其浓度和纯度。将SUVEC细胞接种到6孔培养板上,待细胞 80%~90%融合时,按 LipofectamineTM 2 000说明进行转染,48 h后观察荧光强度。
1.2.5 PLptn Western blot检测 转染后48 h收集细胞,裂解抽提蛋白,样品处理后,进行Western blot检测。首抗为小鼠单克隆抗体Anti-GFP antibody(1∶2 000),二抗为 HRP标记的抗鼠二抗Goat Anti-mouse IgG/HRP(1∶3 000)。
1.2.6 趋化试验 通过检测猪外周血液中的淋巴细胞的迁移来反映Pig Lptn活性,结果以趋化指数表示。
2 结果与分析
2.1 总RNA浓度和纯度 紫外分光光度计测定提取总RNA的浓度和纯度,结果为OD260/OD280=1.84±0.01,OD260/OD230=1.32±0.01,RNA 的浓度为66±0.01 ng/μ L。
2.2 PLptn cDNA的获得 以从健康成年长白猪胸腺组织中提取总RNA反转录得到的cDNA为模板,扩增出PLptn编码基因全长序列,结果见图1。
2.3 测序结果 测序结果经比对软件Bioxm2.6分析,所得序列与GenBank登录号为NM-001134345序列完全一致。
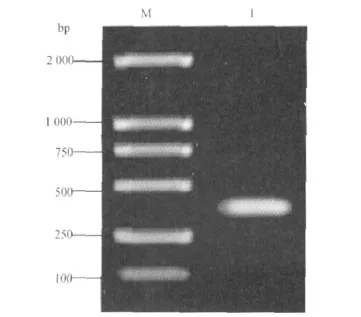
图1 PLptn的PCR扩增
2.4 pEGFP-PLptn转染SUVEC结果 紫外分光光度计测定质粒pEGFP-PLptn和pEGFP-C1的浓度和纯度,结果为 pEGFP-C1:OD260/OD280=1.94±0.01,OD260/OD230=2.27±0.01,浓度为378.8±0.01 ng/μ L;pEGFP-PLptn:OD260/OD280=1.93 ±0.01,OD260/OD230=2.35±0.01,浓度为438.2±0.01 ng/μ L。利用荧光倒置显微镜对转染pEGFP-PLptn质粒的SUVEC观察,发现转染pEGFP-PLptn后48 h荧光表达量最多,见图2-3、2-4,图2-1、2-2为pEGFP-C1转染48 h后结果。
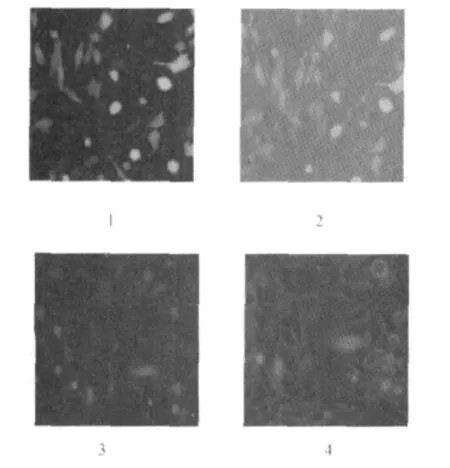
图2 pEGFP-C1和 pEGFP-PLptn转染SUVECs后的荧光显微镜检测(100×)
2.5 PLptn在SUVEC中表达的检测 Western blot结果显示重组载体样品表达约43 ku的蛋白,而空载体pEGFP-C1表达自身约33 ku的蛋白,说明构建的重组质粒pEGFP-PLptn在SUVEC中成功表达,PLptn基因编码的蛋白大约10 ku,与已知相符,结果见图3。
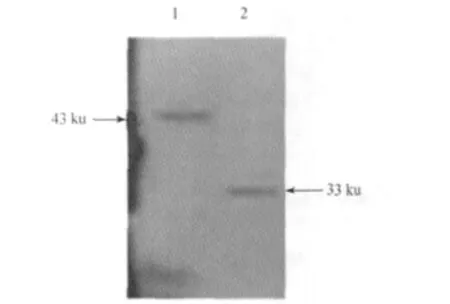
图3 Western blot检测PLptn的表达
2.6 趋化试验结果 用pEGFP-PLptn转染细胞系SUVEC,趋化试验检测融合蛋白对猪外周血淋巴细胞的趋化活性,结果如下:趋化游走距离(A):32 μ m,自发游走距离(B):11 μ m,趋化指数(A/B)为2.9。表明PLptn能在SUVEC中表达,而且表达的PLptn具有趋化猪外周血淋巴细胞的生物学活性。
3 讨论
Lptn在调节免疫系统平衡、增强黏膜免疫、抗肿瘤等方面都发挥重要作用,临床上与肾炎、移植排斥、病毒感染、过敏性哮喘等多种疾病相关[3]。对其功能和作用机制的研究已相对成熟。PLptn编码由110个氨基酸残基组成的多肽,该多肽序列含Chemokine-C 、Chemokine、Chemokine-CC 和 SCY 等功能域,与羊和人的氨基酸序列相似率分别为81.6%和68.4%[4]。具有相似序列的蛋白质具有相似的功能,又PLptn具有与Lptn相同的结构域Chemokine-C,这提示我们PLptn可能具有上述和人淋巴细胞趋化因子Lptn相似的功能。这为研究PLptn的功能和作用机制提供了一种思路,我们可以参考Lptn的研究方法来研究PLptn的功能。本试验成功克隆了猪淋巴细胞趋化因子编码基因全长序列。构建了pMD-18T-PLptn和真核载体pEGFPPLptn,经鉴定均无误。转染细胞Western blot结果显示,目的基因已成功表达,体外趋化活性试验检测融合蛋白对猪淋巴细胞具有趋化活性,为研究PLptn生物学功能奠定了基础。
[1]Ordway D,Henao-Tamayo M,Smith E,et al.XCL1(lymphotactin)chemokine produced by activated CD8(+)T cells during the chronic stage of infection with My cobacterium tuberculosis negatively affects production of IFN-gamma by CD4(+)T cells and participates in g ranuloma stability[J].Leukoc Biol,2007,82(5):1221-1229.
[2]Zavala-Flores L M,Villatoro-Hernandez J,Gamez-Escobedo A,et al.Production of biologically active human lymphotactin(XCL1)by Lactococcus lactis[J].Biotechnol Lett,2009,31(2):215-220.
[3]Zhu C F,OU Z L.Research Progress of Lymphotactin(Ltn)[J].Foreign Medical Sciences:Section of Pathophysiology and Clinical Medicine,2002,22(4):320-322.
[4]宋豪,张德礼.猪淋巴细胞趋化因子XCL1的电子克隆和编码区序列RT-PCR验证[J].动物医学进展,2008,12:11-16.
[5]Blaschke S,Brandt P,Wessels J T,et al.Expression and Function of the C-Class Chemokine Lymphotactin(XCL1)in Wegener′s G ranulomatosis[J].J Rheumatol,2009,36(11):2491-2500.
[6]Rosas-T araco A G,Higgins D M,Sánchez-Campillo J,et al.Intrapulmonary delivery of XCL1-targeting small interfering RNA in mice chronically infected with Mycobacterium tuberculosis[J].Am J Respir Cell Mol Biol,2009,41(2):136-145.
