三江源区高寒草甸43种植物繁殖体质量比较
2010-05-30王慧春周华坤赵新全刘泽华
柯 君,王慧春,周华坤,赵新全,刘泽华
(1.青海师范大学生命与地理科学学院,青海西宁 810008;2.中国科学院西北高原生物研究所,青海西宁 810001;3.中国科学院西北高原生物研究所 高原生物适应与进化重点实验室,青海西宁 810001)
作为植物生活史中一个明显重要的特征,繁殖体质量很早就被生态学家所重视,早在1942年Salisbury[1]就发现,群落中植物物种之间的繁殖体质量有一个变化范围,且这种变化幅度还非常大,可以跨11个数量级以上。然而,繁殖体质量的种内变异则控制在一个比较小的幅度内,各个植物种的繁殖体质量是一个比较稳定的性状[2]。已有研究表明,种子质量与种子的传播距离、种子库寿命、植物定居成功率、植物繁殖力相关[3-5]。将繁殖体质量作为生物学的一个重要性状来关注,将大大拓展植物群落学和种群生态学的研究内容,促进和深化这些学科的发展;对繁殖体质量的研究,还将促进繁殖生物学、全球变化生物学和动植物相关关系研究等学科的发展。英国、阿根廷、澳大利亚、新西兰等国对大量繁殖体的质量进行了专门的比较研究,并探讨了其生态学意义[4,6]。国内关于不同地域之间和地域内不同群落之间大规模的植物繁殖体质量变化的研究有零星报道[7-9]。
在青海三江源区高寒草甸,草地退化、生物多样性丧失和杂草入侵都很突出[10]。目前关于高寒草甸植物生活史繁殖对策的研究很少,在高海拔三江源区对大量植物物种繁殖体质量的比较研究尤为少见,因此,在一定程度上影响了对高寒草甸生活史对策、物种多样性维持、种群扩散、杂草侵入、生物多样性丧失机制的认识。
本研究选择了位于青海省三江源境内的果洛州玛沁县高寒草甸43种植物(隶属于13科32属),对其繁殖体的质量进行了比较,对研究区植物繁殖体的生存对策进行了探讨,旨在了解高寒草甸植物繁殖体的形态特征、生态功能,为退化草甸植被形成过程和种群变化提供一定的研究基础,并为退化草地的恢复治理和生物多样性保护提供理论基础。
1 材料和方法
1.1 研究区概况 试验所用繁殖体采自青海省果洛州玛沁县境内的军牧场地区,地理位置为34°20′~34°22′N,100°29′~100°30′E,平均海拔4 120 m,范围3 800~4 800 m。该地区为典型的高原大陆性气候特点,无四季之分,仅有冷暖季之别,冷季漫长、干燥而寒冷,暖季短暂、湿润而凉爽。温度年差较小而日差悬殊,太阳辐射强烈。日照充足,日照时数平均在2 500 h以上,年总辐射量623.8~629.9 kJ/cm2。冷季持续时间长达7~8个月,且风大雪多;暖季湿润,长 4~5个月。平均气温在0℃以下,全年无绝对无霜期。年降水量420~560 mm,多集中在 5-10月。土壤为高山草甸土和高山灌丛草甸土,土壤表层有机质含量丰富。矮嵩草草甸为该地区主要的冬春草地。建群种为矮嵩草,主要的伴生种有:小嵩草、二柱头藨草、垂穗披碱草、早熟禾、异针茅、短穗兔耳草、矮火绒草、细叶亚菊、兰石草、美丽风毛菊和甘肃马先蒿等[11]。
1.2 试验方法 供试植物共有43种,均为种子植物,分属于13科。对其中的30种测量果实,13种测量种子。全部繁殖体于2006年秋季采集。本研究分析的是种子、闭果和单种果实。
繁殖体质量的测定方法:取100粒繁殖体,测其风干质量,每种植物重复5次。由于繁殖体附属物对植物的传播具有价值,禾本科植物带芒(如果有芒),菊科植物果实带冠毛,掌叶大黄果实带翅测量。
根据文献[12]引用学名并确定植物的生活型和生态特性。
2 结果与分析
所测定的43种植物的繁殖体(种子或果实)质量差别很大,最小的单粒质量不足0.03 mg,如毛茛;最大的单粒质量超过5 mg,如掌叶大黄。
对传播体为果实的植物,可以大致分为5个组别(表1)。矮火绒草瘦果单粒质量最轻(0.049 mg),掌叶大黄瘦果单粒质量(5.905 mg)最重。对传播体为种子的植物,也可大致分为5个组别(表1)。毛茛种子单粒质量最轻(0.028 mg),马尿泡种子单粒质量最重(3.98 mg)。

表1 繁殖体单粒质量分组
一年生或一、二年生植物皆为杂草,繁殖体质量偏小(平均粒质量0.517 mg),其中以果实作为传播体的仅臭蒿1种,且其果实(单粒质量0.102 mg)最轻,以种子作为传播体的植物有4种,粒质量大小为湿生扁蕾种子(0.136 mg)<甘肃马先蒿种子(0.486 mg)<灰绿藜种子(0.575 mg)<小米草种子(1.285 mg)(图1)。

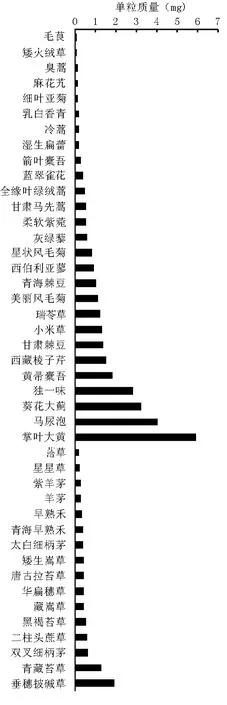
图2 不同生态功能类群植物的繁殖体大小

图3 不同科植物的繁殖体大小
在多年生植物中既有杂草又有牧草,繁殖体平均粒质量0.969 mg,以果实作为传播体的植物有28种,繁殖体平均粒质量为0.948 mg,以种子作为传播体的植物只有9种,繁殖体平均粒质量为1.036 mg。由此可见,多年生植物较一年生植物的繁殖体质量大,不同传播体植物的繁殖体质量差异不大。
从植物的生态功能类群来看,优良牧草的繁殖体质量偏小(平均粒质量0.518 mg),杂草的繁殖体质量偏大(平均粒质量1.122 mg),其中以果实作为繁殖体的杂草平均粒质量为1.214 mg,以种子作为繁殖体的杂草平均粒质量为0.987 mg,两者差异不显著。
禾本科(平均粒质量0.487 mg)和莎草科植物(平均粒质量0.559 mg)的繁殖体质量整体偏小,但个别也有偏大的,如禾本科的垂穗披碱草果实粒质量达1.92 mg,莎草科的青藏苔草果实粒质量为1.273 mg,菊科植物的繁殖体质量有大有小(平均粒质量0.778 mg),如矮火绒草果实粒质量只有 0.049 mg,葵花大蓟果实粒质量高达3.179 mg。
3 讨论
3.1 43种植物繁殖体质量的变化与分布繁殖体质量在植物学和生态学研究上都是一个重要的生物性状。其中种子质量与种子库寿命的关系受到了生态学家的广泛关注。Thompson等[4]在研究了97种英格兰植物后发现,小粒、近圆球形繁殖体易于形成持久繁殖体库;后来有许多研究者[13-14]得出同样的结论。持久繁殖体库有利于植被缓冲空间或时间上不可预测的干扰所造成的破坏[15]。
在一个既定繁殖分配值下,繁殖体大小和繁殖体数量存在权衡关系[16],因此,小粒繁殖体常与大的产种量相伴[4]。与大粒繁殖体植物相比,小粒繁殖体植物有更大的多度范围、更广泛的空间占有量、出现的年份更多[17],易被生物和非生物等传播媒介所携带和传播[18-19],且小粒繁殖体可能至少部分地表现了抵御捕食的功能[3]。
本研究所涉及的43种植物中,繁殖体质量小于1 mg且同时比较接近圆球形的植物有15种:毛茛、麻花艽、湿生扁蕾、兰花翠雀、唐古拉苔草、甘肃马先蒿、臭蒿、细叶亚菊、乳白香青、紫羊茅、羊茅、全缘叶绿绒蒿、二柱头藨草、灰绿藜、西伯利亚蓼,其中多年生植物占 7 3.3%,一年生或一、二年生植物占26.7%,它们可能具有较大的散布能力和持久繁殖体库[20]。如果这些植物具有持久土壤繁殖体库,则意味着它们具有适应频繁干扰的能力。从目前高寒草甸植被的现状看,上述植物在植物群落中的地位比较明显,但它们是否具有持久繁殖体库还有待下一步研究予以验证。
3.2 生活型与繁殖体大小的关系 植物生活型是植物对环境条件适应后在其生理、结构上的一种具体反映[21],是植物生活史的一个重要方面。繁殖体质量与植物生活型可能有关。一般情况下,灌木比草本植物生产的繁殖体大,乔木和藤本比灌木的繁殖体大,可能是植物高度改变繁殖体散布的效率,具有小繁殖体的高植物能形成有效的散布距离[22]。本研究只涉及冷蒿1种小灌木,其余都是草本植物,无法作比较。
以往研究表明,多年生植物平均繁殖体质量要高于一年生植物[23-24],但是也有一些研究表明种子质量与植物体的寿命无关[25]。试验结果进一步证明了多年生草本植物较一年生草本植物的繁殖体质量大(P<0.05)。一个可能的解释是一年生植物要求繁殖体库分担延续生命的危险更要强烈,分担这种危险的方式之一就是产生大量的小繁殖体,来扩大风媒植物或无特别形态结构的繁殖体的时空分布[26];另一个可能的解释是大繁殖体需要更多的发育时间[27];第3个可能的解释是具有较大繁殖体的植物一般具有较低的比叶面积,而常绿的、寿命长久的植物种类一般具有较低的比叶面积,具有较低比叶面积的植物有较低的单位质量的氮含量,既使在有利的条件下也只有缓慢的相对生长率,具有较高比叶面积的植物每光合单位酶具有较多的光捕捉面积,具有较快的潜在相对生长率[28-29]。
大繁殖体更适应严酷的环境[30],可能机理是较大繁殖体在幼苗开始的生长阶段,具有更大比例的繁殖体储藏物供应幼苗结构或应用于呼吸作用以及修复被伤害的部位。本研究结果表明,高寒草甸优良牧草繁殖体质量较杂草的轻;菊科植物的繁殖体质量偏大,而禾本科和莎草科植物的繁殖体质量整体偏小;在草甸植物中,菊科植物属于杂草类,所以二者结果是一致的。这是导致三江源区高寒草甸草地退化、生物多样性丧失和杂草入侵的原因之一。
3.3 繁殖体附属物与其传播、土壤库及发芽对策 繁殖体附属物在植物的传播和定居方面具有重要的作用,如已有人指出:繁殖体通过具翅、柔毛或羽状物实现风媒传播[31]。禾草植物繁殖体可通过芒的主动运动将自身固结在土壤表面的裂隙中[32-33]。本研究中75%的菊科植物具有冠毛;蓼科植物掌叶大黄、玄参科植物小米草有翅,这些植物都能被风传播;89%的禾本科植物具芒,它们可能具有固结自身的作用。至于它们确切的传播方式有待于进一步验证。
有关文献报道[34-35],大且细长繁殖体和具有吸湿芒、冠毛或齿的繁殖体具有迅速萌发的趋势。在本研究的植物中,菊科植物繁殖体偏大且和禾本科植物一样多数繁殖体具有附属物。至于它们是否能够迅速萌发,有待下一步研究予以验证。
试验对高寒草甸主要植物的繁殖体质量做了多物种比较研究,但还应在多物种基础上调查繁殖体传播、土壤繁殖体库及持久性,观测繁殖体发芽对策,以便分析植物的干扰适应机制,为三江源地区的草地管理与退化植被恢复提供一定依据。
[1] Salisbury E J.The Productive Capacity of Plants[M].London:G.Bell and Sons,1942.
[2] Harper J L,Lovell P H,Moore K G.The shapes and sizes of seeds[J].Annual Review of Ecology and Systematics,1970(1):327-356.
[3] Thompson K.Seed and seed banks[J].New Phytologists,1987,106(1):23-34.
[4] Thompson K,Band S R,Hodgson J G.Seed size and shape predict persistence in soil[J].Functional Ecology,1993(7):236-241.
[5] Lioret F,Casanovas C,Penuelas J.Seedling survival of Mediterranean shrubland species in relation to root:shoot ratio,seed size and water and nitrogen use[J].Functional Ecology,1999,13:210-216.
[6] Moles A T,Hodson D W,Webb C J.Seed size and shape and persistence in the soil in the New Zealand flora[J].Oikos,2000,89:541-545.
[7] 仲延凯,包青海,孙维,等.割草干扰对典型草原土壤种子库种子数量与组成的影响——Ⅲ 120种植物种子的大小与重量[J].内蒙古大学学报(自然科学版),2001,32(3):280-286.
[8] 胡发成,于天明,段军红,等.祁连山东部北坡植被垂直分布特征及保护措施[J].草业科学,2007,24(1):13-16.
[9] 刘有军,王继和,马全林,等.甘肃省荒漠种子植物科的区系分析[J].草业科学,2008,25(5):22-27.
[10] 文海燕,傅华,赵哈林.退化沙质草地植物群落物种多样性与土壤肥力的关系[J].草业科学,2008,25(10):6-9.
[11] 陈学林,梁艳,齐威,等.一年生龙胆属植物的繁殖分配及其花大小、数量的权衡关系研究[J].草业学报,2009,18(5):58-66.
[12] 中国科学院西北高原生物研究所青海植物志编辑委员会.青海植物志[M].西宁:青海人民出版社,1997.
[13] Peco B,Traba J,Levasor C.Seed size,shape and persistence in dry Mediterranean grass and scrublands[J].Seed Science Research,2003,13:87-95.
[14] Grime J P.Plant strategies,vegetation processes,and ecosystem properties[M].Chichester:John Wiley&Sons,2001.
[15] Thompson K.The functional ecology of seed banks[A].Fenner M(eds).Seeds:the ecology of regeneration in plant communities[M].Wallinford:CAB International,1992:231-258.
[16] Leishman M R.Does the seed size/number trade off modeldetermine plant community tructure?An assessment of the model mechanisms and their generality[J].Oikos,2001,93:294-302.
[17] Guo Q,Brown J H,Valone T J,et al.Constrains of seed size on plant distribution and abundance[J].Ecology,2000,81:2149-2155.
[18] Venable D L,Brown J S.The selective interactions of dispersal,dormancy,and seed size as adaptations for reducing risk in variable environments[J].A-merican Naturalist,1988,131:360-384.
[19] Greene D F,Johnson E A.Seed mass and dispersal capacity in wind-dispersed diaspores[J].Oikos,1993,76:211-220.
[20] 周兴民,王启基,赵新全.中国嵩草草甸[M].北京:科学出版社,2001.
[21] Muelier D D,Ellenberg H.Aims and methods of vegetation[M].New York:John Wiley&Sons,1974.
[22] Thompson K,Rabinowitz D.Do big plants have big seeds?[J].American Naturalist,1989,133:722-728.
[23] Telenius A,Torstesson P.Seed wings in relation to see size in the genus Spergularia[J].Oikos,1991,61:216-222.
[24] Silvertown J W.Seed size,lifespan and germination date as co-adapted features of plant life history[J].American Naturalist,1981,118:860-864.
[25] Jurado E,Westoby M,Nelson D.Diaspore weight,dispersal,growth form and perenniality of central Australian plants[J].Journal of Ecology,1991,79:811-830.
[26] Brown J S,Venable D L.Evolutionary ecology of seed bank annuals in temporally varying environments[J].American Naturalist,1986,127:31-47.
[27] Baker H G.Seed weight in relation to environmental conditions in California[J].Ecology,1972,53:997-1010.
[28] Garnier E.Growth analysis of congeneric annual and perennials grass species[J].Journal of Ecology,1992,80:665-675.
[29] Reichp P B,Walters M B,Ellsworth D S.Leaf lifespan in relation to leaf,plant,and stand characteristics among diverse ecosystems[J].Ecological Monographs,1992,62:365-392.
[30] Willson M F.Plant Reproductive Ecology[M].New York:Wiley,1983.
[31] Grime J P.Plant Strategies,Vegetation Processes,and Ecosystem Properties[M].Chichester:John Wiley&Sons,2001.
[32] Peart M H.Experiments on the biological significance of the morphology of seed-dispersal units in grasses[J].Journal of Ecology,1979,67:843-863.
[33] Peart M H.The effects of morphology,orientation and position of grass diasporas on seedling survival[J].Journal of Ecology,1984,72:437-453.
[34] Thompson K.Morphology and color[A].Hendry G A F,Grime J P.Methods in Comparative Plant Ecology:A Laboratory Manual[C].London:Chapman&Hall,1993:194-196.
[35] Rees M.Trade-offs among dispersal strategies in the British flora[J].Nature,1993,366:150-152.
