马尾松优树无性系生长、干形和木材密度评价
2010-05-08储德裕刘青华金国庆秦国峰周志春
储德裕,刘青华,金国庆,秦国峰,周志春
(1. 浙江省淳安县姥山林场,浙江 淳安 311700;2. 中国林业科学研究院亚热带林业研究所,浙江 富阳 311400)
马尾松优树无性系生长、干形和木材密度评价
储德裕1,刘青华2,金国庆2,秦国峰2,周志春2
(1. 浙江省淳安县姥山林场,浙江 淳安 311700;2. 中国林业科学研究院亚热带林业研究所,浙江 富阳 311400)
摘要:利用浙江淳安国家马尾松(Pinus massoninna)种质库中22年生来自9个产地(或省区)180个优树无性系的保存林材料,研究马尾松优树无性系生长和材性在产地间和产地内的遗传变异规律,并对保存的优树无性系进行综合评价。结果表明,马尾松优树无性系生长、形质和木材基本密度皆存在显著的产地间和产地内变异,其中产地内无性系间的变异为产地间变异的1.87~2.38倍;与生长性状比较,木材基本密度在产地间所占的变异分量相对较大;除湖南产地优树无性系年轮宽度从髓心向树皮呈单峰变化外,其它8个产地优树无性系的年轮宽度皆随形成层年龄的增加而逐渐变窄;马尾松优树无性系的木材基本密度从髓心至树皮逐渐增大,到达一定年龄后渐趋稳定,但四川和广东两产地优树无性系木材密度的径向均匀性较低。早晚相关分析结果显示,除江西、广东和四川产地优树无性系木材基本密度的早期选择年龄晚于径向生长外,马尾松不同产地优树无性系的木材密度和径向生长皆可在5年生时开始早期选择。通过聚类可将180个优树无性系归为3大类和4个亚类,并综合评选出20个速生优质的优树无性系,其主要集中在第1类和第2类第2亚类。
关键词:马尾松;优树无性系;生长;材性;遗传变异;评价
林木种质资源是树种长期演化的产物,在树种之间、种内不同地理种源间及种源内不同个体之间皆存在着显著的差异[1]。林木种质资源是事关国家和区域可持续发展的重要战略资源。种质资源的保存除了可以保护遗传多样性外,还是林木育种的物质基础。育种工作成效的大小在很大程度上取决于能否全面掌握种质资源并进行合理的利用。在林木集约经营中,由于只注重少数经济性状的选择,林分的遗传基础变窄,抗逆性变弱,为此很多国家加强对林木育种种质的收集和保存以应对未来生产、生活需求和气候的改变而导致育种目标变化,如芬兰、美国等在保护天然基因资源的同时,还设置了优树收集区来保存丰富的种质资源[2]。我国从20世纪80年代初全面开展主要造林树种的遗传改良以来,已收集保存了数万份种源、家系、优树无性系等育种种质,杉木、马尾松和油松等都建立了规模较大的育种种质基因库[3~5],现在其林龄已达到20~30年生,然而却忽视对这些林木育种种质的遗传整理及评价,缺乏种质资源长期保存和有效利用的理论基础。
马尾松(Pinus massoninna)是我国南方山地的主要针叶商品用材树种之一,生长快,适应性强,广泛用于制浆造纸、建筑和松香制造等。自20世纪70年代末我国全面开展马尾松地理种源试验研究以来,现已系统揭示了马尾松生长、形态、物候、材质材性等种源变差和地理遗传变异规律,分别不同造林区筛选出一批优良种源[6~8]。同时,南方各主要产区开展了大规模马尾松优树选择和种子园营建技术研究,全国共选出优树5 500多株,建立初级无性系种子园1 100 hm2[9]。为实现马尾松长期有效遗传改良,中国林业科学研究院亚热带林业研究所于1986年在浙江省淳安县姥山林场林木良种基地收集保存了南方10个省区1 076个优树无性系,建成了国内最大的马尾松一代核心育种群体,现被列为国家马尾松种质资源库。目前该马尾松一代核心育种群体保存完整,林木生长良好。本文利用不同产地和产地内不同无性系材料,系统研究马尾松生长、形质和木材基本密度的产地及产地内无性系的遗传变异,揭示不同产地无性系年轮宽度和木材基本密度的径向变异规律及性状早晚相关性等,并对马尾松一代育种亲本无性系进行综合评价,为更好地实施马尾松育种种质的长期保存和有效利用提供重要理论依据和科学指导。
1 材料与方法
1.1 材料来源
试验材料来自浙江省淳安县姥山林场马尾松一代核心育种群体。该育种群体位于浙江省千岛湖东南湖区,共收集保存了来自10个省区的1 076个优树无性系,其自然概况及立地条件见参考文献[8]。收集区建于1984-1986年,1984年下半年进行林地整理,并开挖水平栽植带,带面宽1.2 m,带间距4.0 m。1985年1月利用当地1年生马尾松大田苗栽植定砧,株距2.5 m,穴规40 cm×40 cm×30 cm。1986年4月下旬至5月中旬采集优树无性系采穗圃母株上部生长健壮的穗条,采用髓心形成层对接法进行嫁接[10],嫁接成活率和保存率均在80%以上。无性系按产地分区栽植,各区内无性系完全随机化排列,3~5次重复,4株小区。嫁接后每年皆采用劈抚措施进行林地管理。此外在嫁接后的第10年和第15年时进行2次弱度间伐,伐除一些生长异常、病虫害严重的植株,目前每小区保存2株分株。
1.2 试验林调查和木材基本密度测定
2008年9月选择安徽(无性系编号3***)、浙江(无性系编号5***)、江西(无性系编号6***)、湖南(无性系编号7***)、福建(无性系编号9***)、广东(无性系编号10***)、广西(无性系编号11***)、贵州(无性系编号12***)、四川(无性系编号13***)9个产地,每产地随机选取20个无性系,测量2分株树高、胸径、树干通直度、树干圆满度(用树干下部3 m高处直径与胸径之比表示)等指标。树干通直度分通直、较通直、一般、弯曲、严重弯曲5级,分别记作5、4、3、2和1。同时在植株胸高处上坡方位用6 mm直径的生长锥钻取一髓心至树皮的完整无疵木芯。对所取木芯自髓心向外每3个年轮切成一段,量测其宽度(Wi),并用最大含水量法测定每年轮段的木材基本密度(Di)[11]。
1.3 数据分析
利用各年轮段宽度测定值估算其所代表的年轮段面积:
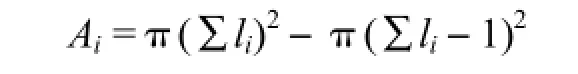
式中,Ai为第i段年轮段面积,li为第i段木芯的长度。
进而计算1~3轮、1~6轮、1~9轮、1~12轮、1~15轮和1~18轮木材的加权平均基本密度BD:

用于试验研究的无性系虽然是按产地分组收集保存的,但因各产地无性系保存所处的立地条件差异较小,可简单地按完全随机化设计进行性状方差分析用以检验产地和产地内无性系效应。方差分析的线性模型为:
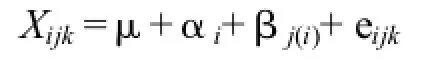
式中,Xijk为第i个产地内第j个无性系的第k个分株的测定值,μ为总平均值,αi为第i个产地的平均值,βj(i)为第i个产地内第j个无性系的平均值,eijk为机误。
方差分析模型见表1。由于产地内各无性系重复次数不等,需计算加权平均重复数n0。产地和产地内无性系重复力分别按以下公式计算:
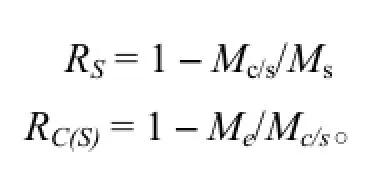
2 结果与分析
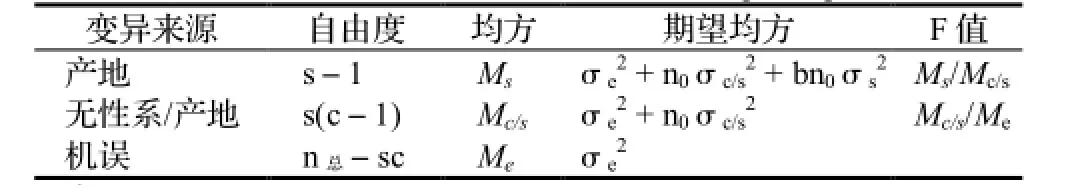
表1 不等重复双因素分组试验方差分析模型Table 1 ANOVA model for double factors unequal repeated trial
2.1 优树无性系生长、形质和木材基本密度在产地间和产地内的遗传变异
表2方差分析结果表明,马尾松一代育种亲本无性系的生长、形质和木材基本密度在产地间和产地内无性系间皆存在显著的遗传差异,其中产地内无性系间的变异大于产地间的变异,前者占总变异的16.13%~38.63%,后者只占总变异的8.46%~23.15%,意味着在充分利用马尾松产地变异的同时还更应注重产地内优树无性系的选择与利用。与胸径、树高和单株材积等生长性状比较,木材基本密度在产地间的变异相对较大,产地选择对于木材密度等材性改良也很重要。由于在马尾松优树选择时皆要求树干通直圆满,因此树干通直度和圆满度在产地间和产地内无性系间的差异相对较小,说明通过优树选择其干形已得到了明显改良。马尾松生长、干形和木材密度的产地重复力都很高,变化在0.80~0.90,生长和木材密度的无性系重复力较高,在0.68~0.79,而树干通直度和圆满度的无性系重复力中等,分别为0.55和0.39。

表2 马尾松优树无性系生长、形质和木材基本密度的方差分析Table 2 ANOVA on growth, form traits and wood density of plus clones
180个来自9个产地的优树无性系其22年生胸径变化在8.43~33.55 cm,平均值为22.0 cm,其中产自广西和湖南的优树无性系胸径生长表现最好,平均胸径皆在24 cm以上,而来自安徽产地的优树无性系胸径生长最差,分别只有广西和湖南产地的79.66%和79.61%。在测定的优树无性系中产地贵州的1208无性系胸径生长量最大,达33.55 cm,是四川产地最小优树无性系1304胸径生长量的3.98倍(表3)。与胸径生长相似,优树无性系树高生长量最大的是广西产地,较树高生长最差的安徽产地平均高出2.90 m左右,但树高生长最好的优树无性系为福建产的9633,达17.87 m。与胸径和树高生长比较,优树无性系单株材积的变幅较大。
尽管不同产地优树无性系树干通直度平均得分值皆在4分以上,但各产地内无性系间差异较大,如来自安徽产地的3413、3608和3410,福建产地的9577、9618和9676,湖南产地的7649和浙江产地的5509和5132等优树无性系干形一般,得分值在3~4分。相对于树干通直度,马尾松优树无性系的树干圆满度在产地间和产地内的变异则较小,变幅分别为0.91~0.95和0.83~0.98,变异系数分别为1.24%和2.59%。来自不同产地的优树无性系其木材基本密度变异系数为4.40%,其中以四川产地优树无性系木材最致密,平均基本密度为0.461 8 g/cm3,广东产地优树无性系木材基本密度最小,仅为0.369 1 g/cm3。然而基本密度在产地内优树无性系间的变异远大于产地间的变异,如福建产地内,9384优树无性系的木材基本密度达0.492 5 g/cm3,而9512优树无性系的木材基本密度仅为0.324 8 g/cm3。

表3 马尾松不同产地优树无性系的生长、形质和木材基本密度Table 3 Growth, form traits and wood density of plus clones from different producing areas
2.2 马尾松不同产地优树无性系年轮宽度和木材基本密度的径向变异及早晚相关
2.2.1 径向变异 从图1可以看出,除湖南产地外,安徽等8个产地优树无性系的年轮宽度皆表现出随着形成层年龄的增加而逐渐变窄的趋势,与作者对马尾松种源试验的研究结果一致[8],如第16至第18轮年轮段的平均宽度为7.62 mm,仅为第1至第3轮的34.71%。来自湖南产地的优树无性系其年轮宽度的径向变异与安徽等产地则略有不同,从髓心向树皮呈单峰变化的趋势,第4至第6年轮段宽度最大(25.93 mm),且第4至第9年轮段的年轮宽度明显宽于其它产地。在径向生长过程中,各产地优树无性系间的平均年轮宽度差异先逐渐增加,在第7至第9年轮段差异最大,变异系数达14.26%,随后其差异不断减小,到第16至第18年轮段时9个产地的平均年轮宽度相差甚小。
图2绘出了9个产地优树无性系木材基本密度的径向变异情况。结果表明,除因松脂等在髓心部位大量沉积导致第1~第3年轮段的木材基本密度测定值异常偏高外,马尾松不同产地优树无性系木材基本密度的径向变异与多数松类和软阔叶树种相似,即木材密度从髓心至树皮逐渐增大,到达一定年龄后渐趋稳定或略有降低[12]。然而不同产地间仍存在一定的差异。在9个产地中,四川和广东产优树无性系的木材基本密度分别为最大和最小,但两产地优树无性系基本密度从髓心向树皮增加幅度较为剧烈,材性径向均匀性较低,而其它7个产地的优树无性系的材性均匀性则较高,在第9轮后木材密度随年龄逐渐趋于稳定。图2结果还显示,除四川产地外,其它8个产地优树无性系间木材基本密度的差异随年龄的增加而逐渐变小。

图1 马尾松不同产地优树无性系年轮宽度径向变异Figure 1 Radial variation of ring width of plus clones from different producing areas
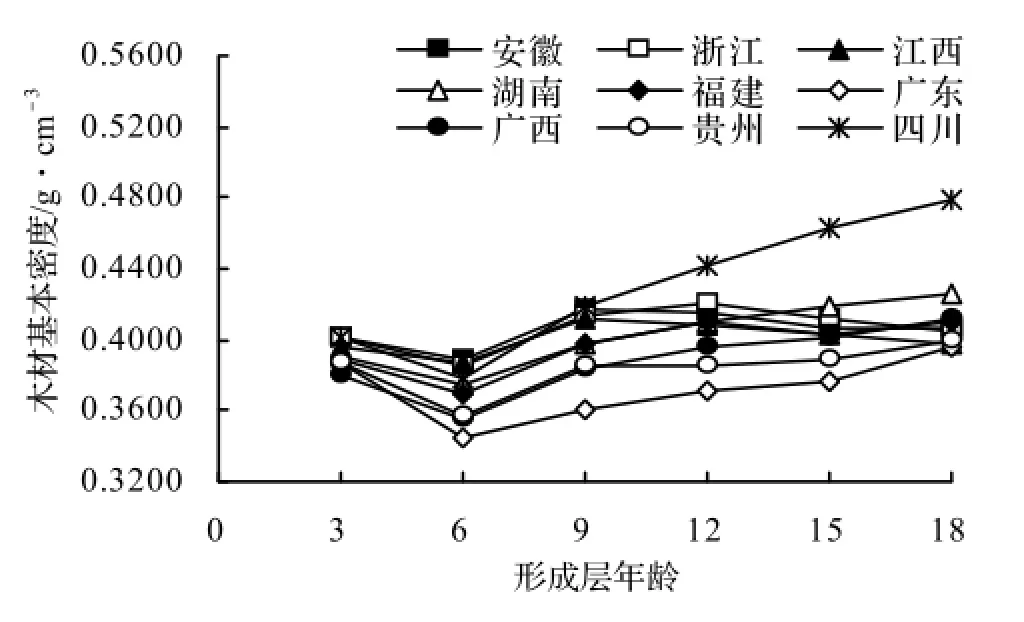
图2 马尾松不同产地优树无性系木材基本密度径向变异Figure 2 Radial variation of wood density of plus clones from different producing areas
2.2.2 早晚相关 表4分别不同产地估算给出了优树无性系年轮宽度和木材基本密度的早晚相关系数。结果显示所有9个产地的优树无性系其第1至第3轮、第1至第6轮、第1至第9轮、第1至第12轮和第1至第15轮的年轮宽度与第1至第18轮的年轮宽度皆呈极显著正相关,且随着年龄的增加,早晚相关系数则稳步增大,表明利用第1至第3轮段(约5年生)的年轮宽度即可对优树无性系生长进行早期选择,而利用第1至第6轮段(约8年生)的年轮宽度进行生长早期选择的效果则更佳。而对于优树无性系的木材基本密度,江西、广东和四川产地除第1至第3轮段与第1至第18轮段的相关性较小外,其它年轮段皆与第1至第18轮段呈高度的正相关,意味着这3个产地优树无性系木材基本密度的早期选择年龄晚于径向生长。其它6个产地优树无性系各年轮段的木材基本密度与第1至第18轮段高度相关,在5年生时可进行早期预测。

表4 马尾松不同产地优树无性系年轮宽度和木材基本密度的早晚相关Table 4 Juvenile-mature correlation of ring width with wood density of plus clones from different producing areas
2.3 马尾松优树无性系聚类分组和种质评价
2.3.1 性状相关 由表5表明,优树无性系生长与干形和木材基本密度的相关性因产地不同而有较大的差异,其中湖南、福建、广西和贵州4个产地优树无性系的胸径和树高与树干圆满度、通直度的相关性较小,生长和干形性状互为独立,而安徽、江西、广东、四川和浙江5个产地优树无性系的胸径或树高与树干圆满度或通直度呈显著正相关,意味着从这5个产地中选择速生的优树无性系其树干形质也可得到一定改良。浙江、江西、湖南和广东产地的优树无性系其胸径或树高与木材基本密度呈高度的负相关,速生的优树无性系其木材基本密度较低,而安徽、福建、广西、贵州和四川产地优树无性系的木材基本密度却受生长的影响较小。若不分产地对所有优树无性系进行性状相关分析,因分析时掩盖了产地效应,未发现优树无性系生长与干形、木材基本密度间存在明显的相关性。

表5 分别产地和不分产地优树无性系的性状相关分析Table 5 Correlation analysis on traits of all tested plus clones and among clones
2.3.2 优树无性系聚类分组 基于胸径、树高、树干通直度、树干圆满度和木材基本密度5个性状的产地均值,对参试产地进行Q型聚类,若以欧氏距离10为阈值,可将9个产地明显地划分为3类:浙江、江西、湖南、广西和贵州5个产地聚为一类,其优树无性系总的特点是生长量较大、干形圆满通直、木材基本密度中等;福建、广东和四川3个产地聚为一类,其特点是生长量中等、干形圆满通直,但3个产地优树无性系的木材基本密度却表现不一,前两个产地的木材基本密度低,而四川产地的木材基本密度高,该类根据基本密度可再分为2个亚类,安徽产地单独聚为一类,其生长、干形相对较差,但木材基本密度较高。
若对参试的180个优树无性系进行直接聚类,同样以欧氏距离10为阈值,可将优树无性系明显地划分为3大类:7606、7631、1137和1208聚为第一类,突出表现为胸径和树高生长量大,22年生时平均值分别达31.69 cm和16.42 m,但木材基本密度略低于另两类;1304和3608两个优树无性系聚为一大类(第三类),其生长量低、干形差,但木材致密;其余174个优树无性系聚为一大类(第二类),其生长和木材基本密度介于第一和第三类之间,干形较为通直。第二类又可以进一步分为四个亚类,第一亚类优树无性系生长相对较快,平均胸径和树高分别为27.83 cm和15.48 m,但木材基本密度较低,平均为0.378 1 g/cm3,其它三个亚类的优树无性系生长相对较慢,木材基本密度相对较大。试验的优树无性系主要集中在第二亚类和第三亚类,各自包括71个和87个无性系。

表6 优树无性系聚类分组及其特性分析Table 6 Cluster analysis and characteristics of all tested plus clones
2.3.3 优良无性系种质选择 这里以生长量(胸径、树高、材积)和木材基本密度皆不低于浙江淳安当地优树无性系均值(代号:51**),树干圆满度不低于0.9,树干通直度得分在4.0以上为约束条件,共筛选出20个生长、形质和材性皆优良的优树无性系(表7)。其中来自广西、贵州和福建产地的优树无性各有2个入选,四川和湖南产地的优树无性系表现较好,各有5个入选,浙江产地的优树无性系仅有1个入选,而江西和广东产地的参试优树无性系皆未入选。入选的优树无性系主要集中在以无性系聚类的第一类和第二类第二亚类,可补充作为马尾松二代育种亲本。

表7 入选马尾松优树无性系的生长、形质和木材基本密度测定值Table 7 Standards of growth, stem traits and wood density for plus clones
3 结论与讨论
研究产地间和产地内生长和材性等主要经济性状的遗传变异分量大小及所占比例对于科学制定林木选择育种策略具有重要的理论指导意义。本文利用马尾松一代核心育种群体中来自9个产地180个优树无性系的保存林材料,发现22年生时马尾松优树无性系生长、形质和木材基本密度在产地间和产地内无性系间皆存在显著的遗传差异,两者重复力较高,进一步证实了马尾松优良种源选择和优良种源内优树选择的重要性与选择潜力。然而不同产地优树无性系间的遗传变异规律却与马尾松全分布地理种源试验结果存在一定的差异[13,8]。如广东、福建两个产地的优树无性系,其胸径和树高平均生长量低于江西、湖南和贵州等中部产地,这与其参试的优树无性系多来自两产地中的次优良种源区有关。因优树选择时未涉及木材密度这一材性指标,其不同产地间的木材基本密度变异规律与种源试验结果基本一致,即表现出南部产地低而北部产地高的趋势[8]。较之于产地间的差异,马尾松生长、形质和材性在产地内优树无性系间差异更为显著,其产地内优树无性系间的变异为产地间变异的1.62~2.66倍。与生长和木材密度比较,优树无性系的树干通直度和圆满度不仅在产地间和产地内的差异性较小,而且在产地内和产地间变异的相对差异也较小,这与马尾松具有树干通直圆满的优良特性及优树干形经过强度选择有关。如各产地无性系树干通直度和圆满度得分分别在4分和0.9以上,这也说明了优树干形选择能很好地达到干形改良效果。
本研究利用生长、形质和木材基本密度5个性状进行聚类,可将马尾松9个产地划分为3类,来自马尾松分布区中部的浙江、江西、湖南、贵州产地和南部的广西产地的欧氏距离较近,聚为一类;南部的福建、广东产地与西部的四川产地聚为一类,其中四川省产地的基本密度较高,而福建和广东产地的基本密度较低,可分别划分两个亚类;北部的安徽产地单独聚为一类,其生长差,但基本密度高。若不分产地,可将所有测试的180个优树无性系归为3大类和4个亚类,其中有174个优树无性系属于第2大类,而速生优质的优树无性系多在其下第2亚类。据张一等利用ISSR分子标记研究发现,当马尾松一代育种亲本间的分子遗传距离处于0.357 ~0.529时,组配亲本间的遗传距离与其子代生长性状具有显著的相关性[14]。因此利用马尾松优树无性系的聚类结果,可为杂交育种亲本的科学选配提供理论指导,以避免杂交育种中亲本选配的盲目性,如可选用湖南、广西等产地中速生的优树无性系与遗传距离较远、木材密度高的四川产地优树无性系开展双亲控制授粉杂交育种,创制生长快、材质致密的新品种。试验综合评选出的速生优质优树无性系,可增补为二代育种的亲本材料,或可用于改良代(1.5代)无性系种子园的营建。
参考文献:
[1] Megraw R A. Wood quality factors in loblolly pine[M]. Georgia:TAPPI Press Atlanta, 1985. 1-88.
[2] 沈熙环. 林木育种学[M]. 北京:中国林业出版社,1990. 6-12.
[3] 郭俊荣,杨培华,谢斌,等. 油松育种资源选择及促进结实技术研究[J]. 西北林学院学报,2004,19(1):46-49.
[4] 王华缄,邓宗富,卢俊豪,等. 杉木基因资源收集、保存和利用的研究[J]. 林业科学,1999,35(5):38-45.
[5] 黄楚光. 马尾松优良种质资源基因库营建策略分析[J]. 福建林业科技,2007,34(3):145-149.
[6] 全国马尾松地理种源试验协作组. 马尾松种源变异及种源区划分的研究[J]. 亚热带林业科技,1987,15(2):81-89.
[7] 周志春,傅玉狮,吴天林. 马尾松生长和材性的地理遗传变异及最优种源区的划定[J]. 林业科学研究,1993,6(5):556-564.
[8] 刘青华,金国庆,张蕊,等. 24年生马尾松生长、形质和木材基本密度种源变异与种源区划[J]. 林业科学,2009,45(10):55-61.
[9] 周志春,秦国峰. 马尾松遗传改良的成就、问题和思考[J]. 林业科学研究,1997,10(4):435-442.
[10] 秦国峰,王培蒂. 马尾松嫩枝嫁接若干基本问题的分析[J]. 浙江林业科技,1992,12(2):1-5.
[11] Smith D M. Maximum moisture content method for determining specific gravity of small wood samples [R]. US Forest Service, Forest Products Laboratory, 1954, Report No: 2014.
[12] Zobel B J,van Buijtenen J P. Wood variation:its causes and control[M]. Berlin:Springer-Verlag,1989. 33-69.
[13] 周志春,秦国峰. 马尾松天然林木材化学组分和浆纸性能的地理模式[J]. 林业科学研究,1995,8(1):1-6.
[14] 张一,储德裕,金国庆,等. 马尾松亲本遗传距离与子代生长性状相关性分析[J]. 林业科学研究,2010(2):215-220.
中图分类号:S781.3
文献标识码:A
文章编号:1001-3776(2010)05-0046-07
收稿日期:2010-04-20;修回日期:2010-06-10
基金项目:“十一五”国家科技支撑专题(2006BAD01A1403),国家林业局林木种质种苗质量监管与保护项目“马尾松种质资源收集保存”
作者简介:储德裕(1964-),男,浙江建德人,工程师,从事林业技术推广和林木良种繁育工作。
Evaluation on Growth, Stem Form and Wood Density of Plus Clones of Pinus massoniana
CHU De-yu1,LIU Qing-hua2,JIN Guo-qing2,QIN Guo-feng2,ZHOU Zhi-chun2
(1. Chun’an Laoshan Forest Farm of Zhejiang, Chun’an 311700, China;2. Research Institute of Subtropical Forestry, CAF Fuyang 311400, China)
Abstract:180 clones from nine producing areas were selected in a 22-year genebank ofPinus massonianafor studying growth and wood properties of plus clones, as well as genetic variation within and among producing areas. And evaluation was made on the utilization value of the preserved resources. The results indicated that growth, stem form and wood density of plus clones had significant differences within and among producing area, and the variation among clones within the same producing area was 1.87-2.38 time higher than that among producing areas. Variance component of wood density among producing areas was larger compared with growth. Ring width of all plus clones decreased from the pith to bark except of Hunan. Wood density of plus clones increased from pith to bark, and was stable at certain age, but that of Sichuan and Guangdong had lower evenness. Juvenile-mature correlation analysis demonstrated that that radial growth and wood density of plus clones could be selected at 5-year except that of Jiangxi, Guangdong and Sichuan. The 180 clones could be divided into three categories and four subclasses by cluster analysis. 20 fast-growing and high quality clones were selected, most of them were in the second category and the second subclass.
Key words:Pinus massoniana; plus-tree clone; growth; wood property; genetic variation; evaluation
