土壤水分对大叶黄杨生长量与光合作用的影响
2010-03-31王志泰
王志泰,包 玉
(贵州大学林学院,贵州 贵阳 550025)
由于中国城市化水平的迅速提高,城市绿化建设的大力发展,城市园林绿化养护用水日益成为城市水资源消耗大户。而全国有2/3的城市常年处于供水不足状态[1]。因此,研究城市园林绿化植物抗旱及耗水规律,发展节水型园林显得尤为重要和紧迫。研究表明,水分(尤其是土壤水分)对植物的生长、蒸腾、光合等生理过程具有明显的影响,从而影响了植物的光能利用效率[2-4]。灌木在城市绿化中常被大量采用,灌木绿地往往栽植密度大,水分需求多,是城市绿地水分管理的重点[5]。近年来,一些学者研究了园林植物耗水规律,通过对树木耗水特性等方面的研究[1,6],选择出耐旱性强、耗水量少的抗旱节水树种,用于城市绿化的空间配置,可以减少其修剪次数,降低养护成本。
大叶黄杨(Euonymus japonicus)又名冬青卫矛,卫矛科卫矛属常绿灌木或小乔木,叶色光亮,嫩叶鲜绿,极耐修剪,为庭院中常见绿篱树种。由于其适应性强,极耐修剪,全国各省普遍栽培。研究表明大叶黄杨对二氧化硫抗性较强,具有很强的滞尘能力,是优良的城市绿化灌木。在发展城市园林绿化和城市生态环境建设方面具有突出的地位;大叶黄杨又可入药,具有祛风温,强筋骨,活血止血功效[7]。
近几年一些学者以大叶黄杨为对象开展了大量的研究,主要包括大叶黄杨的耐荫性[8]、耐寒性[9]、耐旱性[10],大叶黄杨的抗污、吸尘能力[11-13],繁殖栽培[14],病虫害[15]以及光合生理[16-17]等。但是针对不同土壤水分梯度下大叶黄杨生长与光合生理特性变化的研究鲜见报道。本研究通过控制水分试验,探讨不同水分胁迫下,大叶黄杨生长量、叶片脱落与形态变化以及光合作用变化的趋势,寻求控制其生长的临界水分需求规律,为进一步建立园林绿化灌木水分需求动态模型,优化城市绿化管理,科学节水,发展节水型园林提供理论依据。
1 试验材料与方法
1.1 试验材料试验用大叶黄杨苗来自贵阳市花溪区,同龄、同规格,平均苗高42 cm,2008年12月15日将苗木移栽到26 cm口径塑料盆中进行苗木适应性培育,盆内土壤采自贵州大学花溪南校区马尾松林,装盆时种植土与松针土按2∶1混合,基本理化性质为:全氮含量0.93 g/kg,有效氮含量226.9 mg/kg,全磷含量0.57 g/kg,有效磷含量5.4 mg/kg,全钾含量10.54 g/kg,有效钾含量132.5 mg/kg,有机质含量105.7 g/kg,pH值为6.4。每盆种植一株大叶黄杨苗,置于贵州大学花溪南校区林学院教学试验苗圃温室内培养,试验期间不施肥。
1.2 试验处理试验设4个处理:对照(CK),正常供水,土壤含水量一直控制在饱和含水量的90%±5%;轻度胁迫(LS),土壤含水量控制在饱和含水量的70%±5%;中度胁迫(MS),土壤含水量控制在饱和含水量的50%±5%;重度胁迫(HS),土壤含水量控制在饱和含水量的30%±5%,每处理10盆,设3个重复。每天用土壤含水量测定仪(TDR300)结合称量法测量各处理土壤含水量,适时补水,使各处理土壤含水量基本保持恒定。
1.3 测定项目和方法干旱胁迫从移植后的第65天(2009年2月20日)开始。各胁迫处理,按正常浇水的相应比例分别浇水使逐渐达到各处理的设计水平后进行人为控制,用自制苗高测量尺(精度0.1 mm)和游标卡尺分别于胁迫开始的当天,以及以后每周测定苗高和地径,直至第8周,重度胁迫苗木出现严重萎蔫为至。每周记录脱落叶片数。胁迫开始后每隔2周用Li-6400便携式光合分析仪测定同位叶片光合生理指标,测定时间为8:00-10:00,每个处理取顶端向阳方向叶片5~10片进行测定,取平均值。分别测定叶片的净光合速率(Pn)、蒸腾速率(Tr)、光合有效辐射(PAR)、水分利用率(WUE)、光能利用效率(RUE)。于胁迫结束时采摘大叶黄杨苗同位叶,用Epson3490扫描仪扫描后(分辨率300 dpi),按1∶1比例导入AutoCAD软件,测其叶面积、叶长、叶宽,每处理随机测定10片叶,取平均值。

数据用Excel 2003和SPSS13.0处理。
2 结果分析
2.1 土壤水分对大叶黄杨生长量的影响不同土壤水分对大叶黄杨株高生长影响具有显著差异(P<0.01),具体表现为对照和轻度胁迫下株高增长量曲线变化趋势一致,随时间进程逐渐增加。两者之间在各时间进程中亦表现出增长量的差异。中度胁迫和重度胁迫下株高增长量变化曲线基本相似,总体呈现下降趋势,中度胁迫表现出近似线型下降,重度胁迫从胁迫开始的第2周株高增长迅速下降,第2周到第5周呈线型逐渐下降,到第5周株高增长基本停止(图1)。
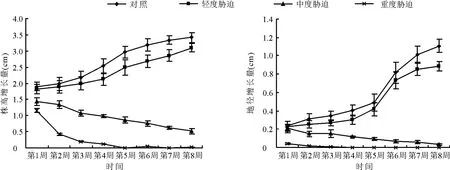
图1 土壤水分对大叶黄杨株高增长和地径增长的影响
不同水分对大叶黄杨地径生长影响的差异性显著。图1结果表明,轻度胁迫和对照的变化曲线趋同,随时间进程呈现出逐渐增加趋势,两者之间在胁迫第2周开始至第6周,地径增长量表现出显著差异(P<0.05),第 6周后差异极显著(P<0.01),胁迫全程轻度胁迫的地径增长量小于对照,而中度胁迫和重度胁迫地径增长量随胁迫进程呈逐渐下降趋势,两者在胁迫伊始至第7周差异极显著(P<0.01),即重度胁迫地径增长量小于中度胁迫,在第8周测定结果趋于相同。另外,还可以从图1中看出,第1周测定结果中,轻度胁迫和中度胁迫的地径增长量与对照的几近相同。
2.2 土壤水分对大叶黄杨叶片的影响大叶黄杨作为园林绿化材料,在控制其生长量的同时,不能忽视其观赏性。本次试验以叶片脱落和叶形变化作为参考指标进行研究。
2.2.1水分对大叶黄杨叶片脱落影响 叶片脱落是植物受到干旱胁迫时的一种普遍应对策略。水分对大叶黄杨叶片脱落的影响以中度胁迫为界,出现两种情况,其一为轻度胁迫对叶片脱落几乎无影响;而中度胁迫下叶片有脱落现象,随胁迫加强,脱落数增加,但总体看叶片脱落数不多,第8周平均脱落数为3.4;其二为重度胁迫叶片脱落现象明显,整个过程脱落数近于中度胁迫的两倍。两者进程各阶段叶片脱落数亦存在极显著差异(P<0.01)。通过日常观察,重度胁迫下,大叶黄杨叶片脱落较多,尤其到胁迫后期,植株濒于死亡,叶片稀少,不利于观赏,如果当做地被植物密植,无法遮蔽黄土。中度胁迫叶片虽有脱落,但与新发叶基本保持平衡,对其观赏价值几乎无影响(表1)。
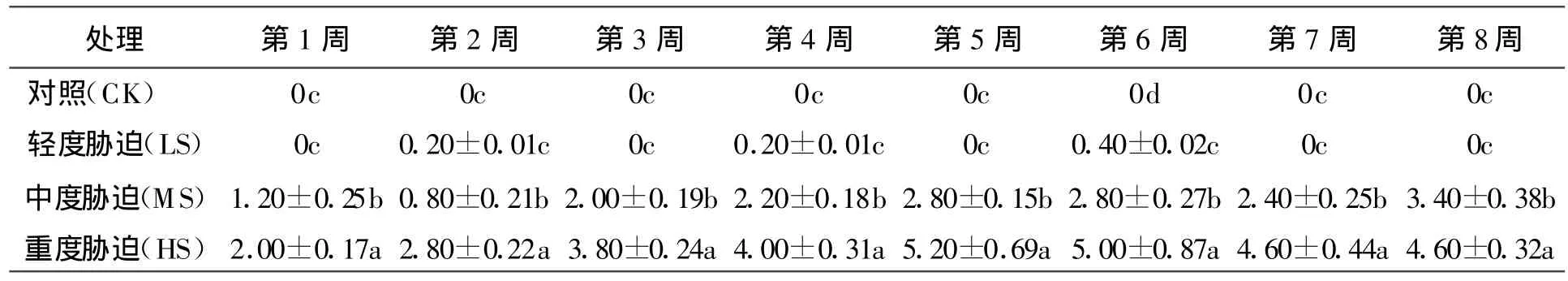
表1 水分对大叶黄杨叶片平均脱落数的影响 片
2.2.2土壤水分对大叶黄杨叶形的影响 不同土壤水分胁迫对大叶黄杨叶形变化有影响,轻度胁迫和对照之间存在显著差异(P<0.05),但差值不大,中度胁迫和重度胁迫与对照差异极显著(P<0.01),且差值较大,而中度胁迫和重度胁迫之间也存在明显差异(P<0.01),但差值不大(表2)。从叶面积差异来看,随着胁迫水平的加强,大叶黄杨叶片变小,中度胁迫下平均叶面积为8.39 cm2是对照平均叶面积15.45 cm2的54.30%。另外,通过表2的叶形相关指标可以看出,叶片形状由长条形向卵圆形变化。
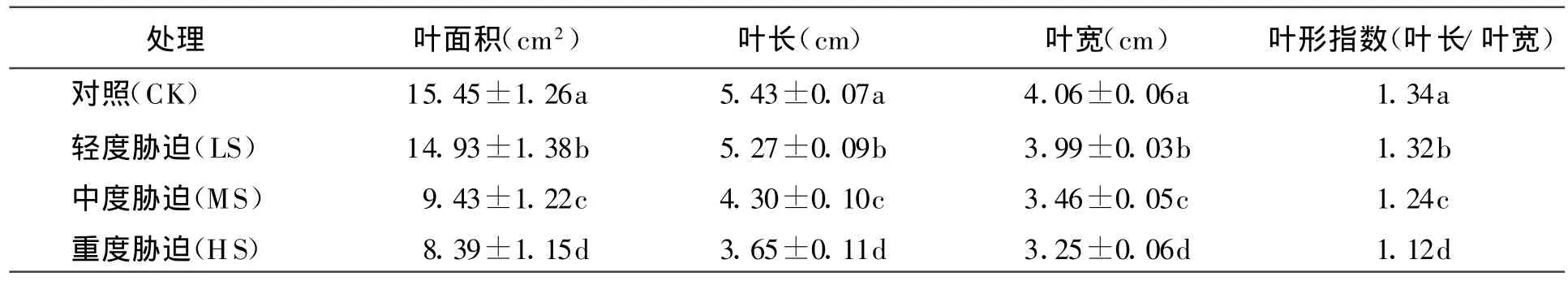
表2 水分对大叶黄杨叶形的影响
2.3 土壤水分对大叶黄杨光合作用的影响
2.3.1土壤水分对大叶黄杨净光合速率和叶片蒸腾速率的影响 不同水分胁迫下,大叶黄杨净光合速率与对照相比有显著差异(P<0.01)(图2)。轻度胁迫下,大叶黄杨前期(第0-4周)光合速率稍低于对照,但趋于平稳,第4周到第6周大幅下降,后期(第6-8周)又趋平稳。中度胁迫和重度胁迫各阶段光合速度明显低于对照,中度胁迫下大叶黄杨净光合速率下降趋势在整个胁迫期逐渐减缓。重度胁迫下的净光合速率下降趋势表现为急-缓-急的趋势(图2)。说明大叶黄杨对土壤水分胁迫较为敏感,并随着胁迫程度的加强,其光合机构受破坏程度加大,致使各胁迫水平下的净光合速率有明显的差异。
轻度胁迫下,大叶黄杨叶片蒸腾速率在前期(第0-4周)没有受到影响,从第4周开始下降,第8周与对照相比存在极显著差异(P<0.01);中度胁迫下,大叶黄杨叶片蒸腾速率随胁迫进程而下降,并逐渐拉大与对照的差距。重度胁迫对大叶黄杨蒸腾速率的影响非常明显,但前期(第0-4周)下降较快,第4周后蒸腾速率基本平稳,第8周接近中度胁迫水平(图2)。
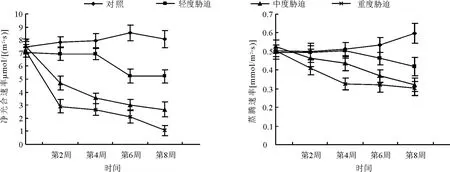
图2 土壤水分对大叶黄杨净光合速率(Pn)和蒸腾速率(Tr)的影响
2.3.2土壤水分对大叶黄杨水分利用效率和光能利用效率的影响 大叶黄杨水分利用效率受不同干旱处理的影响差异较大(图3)。轻度胁迫下,大叶黄杨的水分利用效率前期受到影响较小,与对照差别不大,从第4周开始下降,第6周与对照相比存在极显著差异(P<0.01);中度胁迫下,大叶黄杨的水分利用效率自胁迫开始明显下降,与对照形成极显著差异(P<0.01),到第4周达到最低水平,之后基本保持平稳,并略有上升。重度胁迫时,前期第0-2周水分利用效率显著下降,之后略有上升,到第4周与中度胁迫基本相同,从第4周起又呈现下降趋势,到第8周达到最低水平。从整个胁迫进程来看,总体上水分利用效率表现为:CK>LS>MS>HS(图3)。
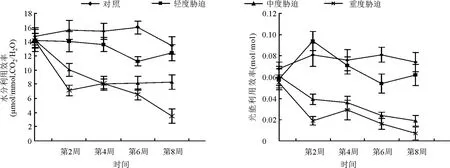
图3 土壤水分对大叶黄杨水分利用效率(WUE)和光能利用效率(RUE)的影响
不同水分梯度对大叶黄杨RUE影响具有极显著差异(P<0.01),具体表现为对照和轻度胁迫RUE曲线变化趋势一致,随时间进程逐渐增加,两者之间在各时间进程中亦表现出RUE的差异。中度胁迫下,大叶黄杨RUE一直呈下降趋势,逐渐拉大了与CK的差距。重度胁迫下,大叶黄杨RUE变化趋势为降-升-降,2周以前迅速下降,2周后又开始上升,到4周后又开始下降,直至胁迫结束(图3)。
3 讨论与结论
3.1 水分胁迫对大叶黄杨的生长量有显著影响水分胁迫对植物生理特性的影响已有大量的研究报道[18]。植物在干旱胁迫下,体内细胞在结构、生理及生化上发生一系列适应性改变后,最终要在植株的生长状况和形态特征上有所表现[19]。本试验通过对这一现象的模拟旨在找到土壤水分梯度与大叶黄杨生长量之间的关系。结果表明,不同的土壤水分梯度下,大叶黄杨株高生长和地径生长存在明显的梯级差异。灌木被用作城市绿化大面积种植做图案或地被植物时,其观赏特征和绿化管理的要求是保持图案的稳定性,园艺管理上常通过修剪来达到这一目的。本试验结果表明可以通过科学的控水使大叶黄杨既达到控制生长量的要求,又可节约水资源和人力资源。
3.2 不同的土壤含水量条件下,大叶黄杨的叶片形态与数量发生了较大的变化当受到干旱胁迫时,作为一种生存策略,植物的叶片形态会发生相应的变化,一般小的表面积/体积比,可以最大限度地减少水分丧失[20]。本试验中,大叶黄杨随水分胁迫加重,其叶形由长条形向卵圆形变化,叶面积减小,叶片数量减少。大叶黄杨常用作绿篱和大面积片植做图案,因此其单株的观赏要求不高,即叶形变化不影响其园林绿化观赏效果,但对其叶片数量有要求。适当的落叶,只要不影响整体的覆盖度,在城市绿化管理中都是允许的。本试验表明在重度胁迫下,大叶黄杨叶片脱落现象明显,因此可以由叶片脱落数和生长量作为确定水分控制临界值的主要指标。
3.3 干旱胁迫抑制了大叶黄杨的光合作用研究报道指出,中度和严重水分胁迫限制植物的生长,导致光合速率降低,WUE升高[21-22]。本试验结果表明,在轻度干旱胁迫的前期,大叶黄杨的光合作用受到了影响,但程度不十分严重,而在中期有明显的下降,后期基本维持不变,说明轻度干旱胁迫对大叶黄杨的生长有一定抑制作用。但本试验中WUE在不同水平的胁迫下均下降,这说明WUE在干旱胁迫下或升或降因植物而宜。本试验用光量子效率法测定了不同土壤水分胁迫下大叶黄杨叶片的瞬时RUE,多数学者用此法研究了农作物的RUE[24-25],本试验的结果表明轻度胁迫下,大叶黄杨的RUE变化趋势与对照一致,这与大叶黄杨在轻度胁迫下植株高生长趋势一致。
3.4 土壤水分中度胁迫可以有效地控制大叶黄杨生长期的生长量宋海鹏等[26]对5种景天属植物通过抗旱性研究结果表明,在园林绿化中可以通过减少灌水量来控制植物的生长,又不影响其观赏效果,本试验与此结果一致。因此,可以初步得出结论:大多数园林绿化灌木,可以通过适度的水分胁迫达到控制生长量、节约灌溉水资源的绿化管理目标。城市绿化的节水灌溉是城市绿化管理的重要内容和新课题,为科学合理地确定城市绿化灌木的节水灌溉策略,还需对大量的园林树种抗旱性比较研究及抗旱生理生态机理的研究来充实其理论依据。
[1]朱妍,李吉跃,史剑波.北京六个绿化树种盆栽蒸腾耗水量的比较研究[J].北京林业大学学报,2006,28(1):66-70.
[2]贾宏涛,赵成义,盛钰,等.干旱地区紫花苜蓿光合日变化规律研究[J].草业科学,2009,26(7):51-60.
[3]代微然,任健,毕玉芬.干旱对假俭草光响应曲线的影响[J].草业学报,2010,19(3):251-254.
[4]李芳兰,朱林海,包维楷.环境胁迫对岷江干旱河谷豆科灌木幼苗根系生长及其结瘤的影响[J].应用生态学报,2009,20(8):1825-1831.
[5]李丽萍,马履一,王瑞辉.北京市3种园林绿化灌木树种的耗水特性[J].中南林业科技大学学报,2007,27(2):44-47.
[6]王瑞辉,马履一.北京15种园林树木耗水性的比较研究[J].中南林业科技大学学报,2009,29(4):16-20.
[7]陈有民.园林树木学[M].北京:中国林业出版社,1990.
[8]包玉,王志泰.不同遮阴处理大叶黄杨叶片生理响应[J].安徽农业科学,2009,37(12):5470-5471.
[9]关雪莲,马清水,张宇,等.大叶黄杨(Euonymus japonicus)和扶芳藤(Euonymus fortunei)耐寒生理指标的变化[J].北京农学院学报,2006,21(3):29-33.
[10]李淑英,周连第,兰彦平.几种常绿阔叶树在北方地区抗旱适应性评价[J].北京农学院学报,2005,20(4):12-16.
[11]杜敏华,张乃群,李玉英,等.大气污染对城市绿化植物叶片叶绿素含量的影响[J].中国环境监测,2007,23(2):86-88.
[12]李瑞雪,张明军,张永芳.石家庄大叶黄杨叶片滞尘量及滞尘颗粒物的粒度[J].城市环境与城市生态,2009,22(1):15-19.
[13]陶玲,任珺,杜忠,等.SO2对兰州市主要绿化树种形态症状的影响[J].环境科学与技术,2009,32(6):34-37.
[14]史红文,杨兰芳,丁昭全,等.污泥和泥炭作基质对大叶黄杨和石楠的应用效果[J].林业科技开发,2009(7):47-48.
[15]江建国,张文颖,白涛,等.10种杀菌剂对大叶黄杨褐斑病田间防治试验[J].中国森林病虫,2009,28(4):30-32.
[16]庄猛,姜卫兵,花国平,等.金边黄杨与大叶黄杨光合特性的比较[J].植物生理学通讯,2006,42(1):39-42.
[17]王勇,杨培岭,任树梅.两种抗旱剂对大叶黄杨光合特性的耦合效应[J].应用生态学报,2007,18(3):514-518.
[18]韩建秋,王秀峰,张志国.土壤水分梯度对白三叶(Tri folium repens)光合作用和根系分布的影响[J].生态学报,2008,28(2):493-499.
[19]季孔庶,孙志勇,方彦.林木抗旱性研究进展[J].南京林业大学学报(自然科学版),2006,30(6):123-128.
[20]任庆成,杨铁钊,刘培玉,等.植物抗旱性研究进展[J].中国农学通报,2009,25(15):76-79.
[21]黎燕琼,郑绍伟,陈泓,等.林木抗旱性研究及其进展[J].世界林业研究,2007,20(1):10-16.
[22]Morgan J A,Legain D R,Mosier A R,et al.Elevated CO2enhances water relations and productivity and affects gas exchange in C3and C4grasses of the Colorado short grass steppe[J].Global Change Biology,2001(7):451-466.
[23]郭慧,吕长平,郑智,等.园林植物抗旱性研究进展[J].安徽农学通报,2009,15(7):53-57.
[24]房全孝,陈雨海,李全起,等.土壤水分对冬小麦生长后期光能利用及水分利用效率的影响[J],作物学报,2006,32(6):861-866.
[25]赵育民,牛树奎,王军邦,等.植被光能利用率研究进展[J].生态学杂志,2007,26(9):1471-1477.
[26]宋海鹏,刘君,李秀玲,等.干旱胁迫对5种景天属植物生理指标的影响[J].草业科学,2010,27(1):11-15.
