植物铁蛋白的研究进展
2010-03-28陈丽萍张丽静傅华
陈丽萍,张丽静,傅华
(农业部草地农业生态系统学重点开放实验室 兰州大学草地农业科技学院,甘肃 兰州730020)
铁是几乎所有生物体所必需的元素。铁含量在地壳中居第4位,但在生物体中的含量则很低。这与Fe3+易形成难溶的化合物导致生物对铁的吸收和利用程度降低有关。虽然土壤中含有大量的铁,但在干旱和半干旱的石灰性土壤上生长的双子叶植物和非禾本科单子叶植物却经常表现出缺铁症状,铁营养失调成为限制植物正常生长发育的重要因素之一[1]。植物缺铁,可引起叶绿素合成受阻,从而导致叶片黄化,光合作用降低,最终造成产量和品质的重大损失,因此,改善植物贮存铁的能力,提高植物的铁营养成分,是解决这个问题的有效途径[2]。铁缺乏时也会影响动物的生长发育,甚至导致一些缺乏铁的综合症,如毛皮动物等对铁缺乏特别敏感,尤其是新生仔兽,易患原发性缺铁病,导致缺铁性贫血[3]。而且对人类来说,铁缺乏是当今世界最为严重的营养缺乏症之一,也是引致贫血的主要原因之一[4],所以提高植物体内铁含量,可为植物自身解除许多由缺铁引起的病症,也可为动物和人类带来福音。
铁蛋白(ferritin)是一种专门存储铁的蛋白质,它广泛存在于动物、植物和微生物体内,既可以储存大量的可被植物利用的铁,又能抵抗环境胁迫[5]。植物铁蛋白储存铁的能力高于其他蛋白[6],且铁蛋白由特殊酶亚基形成,无需导入任何其他基因去诱导产生翻译产物[7]。它是一类多聚体蛋白,在细胞内具有调控铁生物功能的作用,主要在种子形成、叶片衰老或环境铁过量时积累铁,在种子萌发或质体绿化等过程中释放铁,从而调节植物对铁的吸收和释放,维持铁的动态平衡,具有铁储存和避免铁毒害的双重功能[8,9]。在植物体中作为一种胁迫反应蛋白[10],当植物受到寒冷、干旱、强光照、重金属离子等外界环境的胁迫时,植物受到损伤,体内都发现有铁蛋白的存在[11];在铁供应过量的情况下,植物体内其含量是铁正常供应的40~50倍[12]。
近年来,铁蛋白基因作为提高生物体内铁含量,以抵抗外界环境胁迫、预防生物体缺铁性病症的发生而备受关注。自1963年Hyde等[13]第一次报道铁蛋白以来,植物铁蛋白在许多植物中得到证实,如豌豆(Pisum sativum)、大豆(Glycine max)、玉米(Zea mays)、紫花苜蓿(Medicagosativa)等[14-17]。植物体内有多种铁蛋白基因,一些是组成性表达,而另一些则受多种因素诱导,如脱落酸(ABA)、铁离子等[18]。随着生物技术的发展,转基因技术已经成为生物遗传改良的有效途径,利用转基因技术将外源铁蛋白基因转入植物体内,提高植物特别是粮食作物、果树等中的铁含量及抗重金属胁迫能力等的研究也取得了突破性的进展[19-21],不仅可以满足人类对铁的需求,缓解或防御由于铁缺乏而引起的一系列疾病,而且能提高植物对环境的耐受性,在生物治疗中具有重要意义。本研究对铁蛋白的结构、功能及铁蛋白基因的克隆、表达和遗传转化研究等进行了系统总结,旨在为今后进一步的深入研究提供参考。
1 铁蛋白的结构及其分布
1.1 铁蛋白的结构
作为一种铁结合和贮存蛋白,铁蛋白是一类多聚体蛋白。动物和植物的铁蛋白起源于共同的祖先,是一个高度保守的蛋白[6],然而它们的基因结构截然不同。虽然铁蛋白在动物、植物和微生物中基因的内含子个数和所在位置是严格保守的,但是它们中的这些参数是不同的。动物铁蛋白中,内含子的位置和蛋白的二级结构域相关[22,23],而植物铁蛋白的二级结构域与内含子/外显子边界没有明显的关系。植物铁蛋白与动物铁蛋白的同源性为39%~49%,但和细菌铁蛋白没有明显的同源性[5]。
植物铁蛋白是由24个同源或异源亚基所结合成的1个蛋白质复合体,每个亚基包括1个由4股长螺旋组成的束、第5个短的螺旋和1个长的突出环,形成1个中空的蛋白外壳,直径为12~13 nm,壳内含1个复合体状的无机铁核,由细胞中过量铁聚合形成,内径7~8 nm。每分子铁蛋白能以可溶、无毒和生物体可利用的形式储存0~4 500个铁原子不等[6]。所有铁蛋白是由1个球形蛋白质外笼围绕水合铁氧化物而组成,铁原子能够在单一铁蛋白这种铁-蛋白复合物中浓缩[24]。铁蛋白外壳的亚基以4重、3重和2重对称的形式相互排列[25]。在4重与3重轴线上具有离子通道,其中3重轴线附近通道是由亲水残基组成,可结合金属离子如Cd2+、Zn2+、Tb3+或Ca2+等。3重通道是铁的主要通道和Fe2+氧化作用位点,铁离子通过此通道进入核中,在植物和哺乳动物铁蛋白中非常保守。4重对称轴亚单位之间的离子通道比3重轴线上的要窄一些,也是亲水性的,但此结构在动物铁蛋白中却是疏水性的,因此认为此结构是动物和植物铁蛋白的主要区别之一[14]。
1.2 植物铁蛋白的分布
最早通过电子显微镜技术,仅在质体的基质中检测到铁蛋白的存在。尽管在叶绿体中也有铁蛋白,但主要还是分布在低光合活性的非绿色质体如前质体、白色质体、有色体、造粉体以及种子、幼苗、根的顶部[26]和豆科植物年幼的根瘤等特异组织中[15],而有光合活性的叶绿体中却只有少量分布。另外,在植物导管细胞、维管形成层、生殖细胞和衰老的细胞中也发现有铁蛋白的存在[11]。
叶霞等[27]以转菜豆铁蛋白基因的嘎拉苹果(Malus domestica)4个株系为试管苗材料,发现内源铁蛋白和外源铁蛋白基因mRNA的含量均在试管苗的根部表达量最高,茎次之,叶片中最低,进一步在分子水平上证实了铁蛋白的分布规律。
当处于胁迫条件下时,铁蛋白的分布会表现出异常现象。例如,体外培养的大豆细胞中,在质体的外面也发现有铁蛋白的存在。Roswitha等[28]用豌豆突变体dgl和brz发现铁过量时,铁蛋白主要存在于突变体基部叶片的叶绿体中,以及细胞质和质膜与细胞壁的空间。可能是由于过量铁胁迫下,破坏了质膜的完整性,引起邻近坏死细胞中的铁蛋白从衰退的叶绿体中流出所造成的。用硝酸盐培养或者处于黑暗中的菜豆(Phaseolus vulgaris)和豌豆,在其细胞质和没被感染的组织间隙的造粉体,以及薄壁组织细胞中,均发现铁蛋白的存在[29]。这可能是铁蛋白作为一种胁迫反应蛋白,在胁迫条件下产生的一种反应。
2 铁蛋白的功能
2.1 调控铁生物功能、维持铁平衡
生物体内铁的氧化还原反应涉及电子传递过程,参与重要的生化反应如光合作用和呼吸作用[30-32],铁还是多种酶的辅助因子,参加了许多基本的反应如DNA合成、植物固N作用及植物激素合成[17],因此,缺铁会造成严重的营养紊乱,从而影响植物的生理代谢[33]。铁与H2O2反应产生羟自由基,对植物产生巨大的毒性而损害生物有机体,控制细胞中自由Fe2+对植物防御由氧化胁迫带来的损害非常重要[34]。铁蛋白具有强大的贮铁功能,将高毒性的Fe2+转化为无毒的Fe3+螯合物形式,并通过控制细胞中游离Fe2+浓度防御氧化胁迫对植物带来的损害保持植物体内铁的平衡[35],为依赖于铁的生理生化过程起暂时性的铁缓冲作用。
2.2 在植物发育过程中的作用
植物铁蛋白在植物体种子形成、叶片衰老或环境中铁的过量积累方面有着重要功能,它可以在种子萌发或质体绿化过程中释放铁,从而调节植物对铁的吸收和释放[30]。Lobreaux和Briat[32]1991年研究发现在豌豆种子中含有大量的铁,在铁蛋白中贮存的铁占种子总铁含量的92%,在种子生长发育阶段对铁的吸收大量增加,免疫鉴定豌豆成熟种子中也存在铁蛋白,保存的干种子中仍然存在。而种子发芽时,铁蛋白开始降解,产生的铁被用于种苗的生长需要,这些都说明铁蛋白中铁含量在植物发育过程中起重要的作用[36]。在处于发育阶段的玉米叶片中,铁蛋白存在于最幼嫩的叶段和含有衰老细胞的顶端,而富含成熟叶绿体和烯醇丙酮酸磷酸羧基酶的叶片中心部分则不存在铁蛋白[37]。可能是叶片铁蛋白的含量与叶绿体的分化相关,是铁蛋白在植物生长过程中的一种合成调控的途径。
2.3 铁蛋白与非生物胁迫
铁蛋白的积累受各种环境信号的诱导。在寒冷、干旱、机械损伤、衰老、强光照等各种胁迫和重金属、铁过量,烟碱、H2O2、脱落酸、乙烯等化学物质处理的条件下都发现植物中铁蛋白基因的转录量增加数倍,并有大量的铁蛋白存在,另外,抗坏血酸也影响或参与植物铁蛋白的合成[20,32]。当植物处于不利的环境下时,氧化胁迫即占主要地位,抗氧化作用的防御能力减弱,由铁介导的自由基的产生增强,导致代谢失调、脂肪过氧化、蛋白质分解和DNA损伤[10]。由于铁蛋白可容纳大量铁,并以稳定的形式储存,所以对植物抵抗氧化胁迫以及提高植物自身的耐受性等方面有重要作用。
2.4 铁蛋白与生物胁迫
植物中大量积累铁蛋白,还可以对一些真菌的感染、病毒引起的坏死等表现出抗性,保护细胞免受因各种环境胁迫而导致的细胞氧化性损伤[17]。在有病害的植物组织如病毒感染和肿瘤等中,都发现有铁蛋白的积累。如当孢囊线虫(Heterodere glycines)浸染使根瘤的发育和功能受损后,大豆根部即有铁蛋白的积累。此外,病毒感染和肿瘤也促使铁蛋白积累[33]。铁蛋白通过螯合被感染组织和裂解的组织中过量的铁,可以避免铁毒害,同时也能阻止病原体扩散到其他组织,而大豆根部被线虫感染而引起的铁蛋白合成,也可能是大豆结瘤过程被抑制所致[38]。
2.5 抗氧化功能
所有的铁蛋白都可以在有氧条件下与溶液中的二价铁离子反应,铁离子螯合在内部的空心结构中,抑制铁氧化反应,从而保护细胞不受铁过量引起的氧化损害;而且亚铁氧化中心能利用Fenton反应的反应物阻止自由基的产生,所以认为铁蛋白具有抗氧化功能[39]。
在正常的生长条件下,植物铁蛋白天然积累在一些低光合活性的组织中,它们主要在植物的发育过程和植物对环境胁迫的适应性中起作用。在逆境胁迫条件下,植物光合作用中产生的氧自由基及金属离子(主要是铁离子)催化的Fenton反应是氧自由基的主要来源,植物铁蛋白通过贮藏过量的铁,降低植物体细胞内自由铁离子浓度,从而减少氧自由基的产生,降低氧自由基带来的损害[1]。
3 植物铁蛋白基因的克隆、表达调控及转化
3.1 植物铁蛋白基因的克隆
Hyde等[13]于1963年第1次报道铁蛋白以来,植物铁蛋白的研究取得了迅速发展,目前国内外已分别从多种植物中克隆出铁蛋白基因的完整cDNA序列[40-45](表1)。
3.2 植物铁蛋白基因的表达调控
铁蛋白在动物体中定位于胞质中,而在植物体定位在质体中[30]。在单子叶和双子叶植物中已经克隆到一些编码铁蛋白的cDNA序列。Van Wuytswinkel和Briat[46]认为植物铁蛋白亚基首先合成一种前体物质,其N-端具有超过70个氨基酸残基的独特序列。它由两部分组成,第一部分约为40个氨基酸残基的运输肽(transit peptide,TP),负责质体定位,并在进入细胞器后被剪切掉了;另一部分为突出肽(extension peptide,EP)序列,是植物特有的序列,约有30个氨基酸残基,位于成熟铁蛋白N-端,可能是体外蛋白质稳定性调控的一个重要决定因素。植物铁蛋白的突出肽,含有一段α-螺旋,位于蛋白质外壳的外表面,靠近可能参与铁蛋白和其环境之间进行铁交换的通道。在体外铁交换的过程中,这个突出肽是自由基裂解的位点,并且在体内可能导致铁蛋白的降解。突变体分析确认突出肽在维持铁蛋白的蛋白质稳定性中有作用[47]。
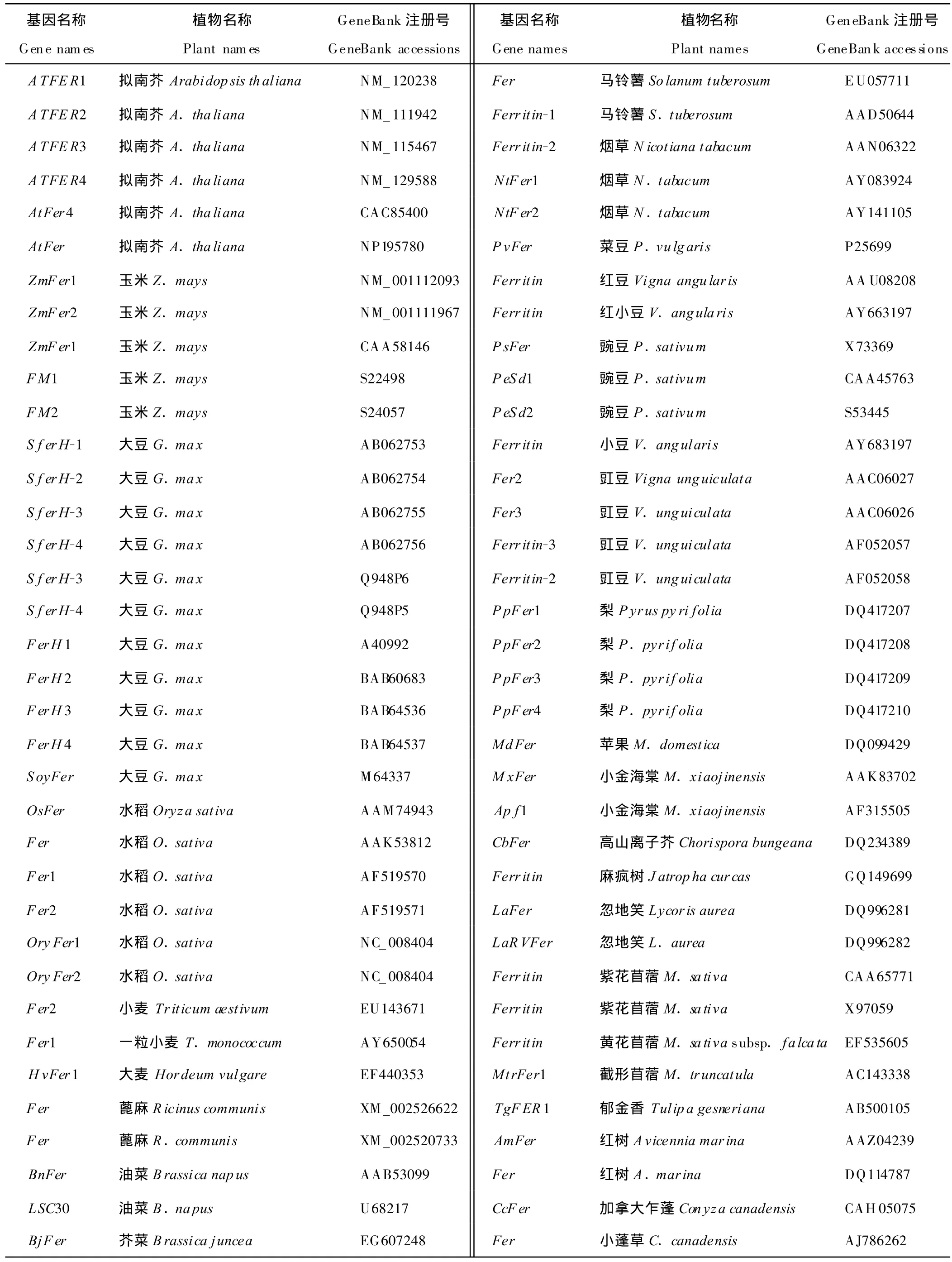
表1 植物中已注册的铁蛋白基因[40-45]Table 1 Gene of ferritin in plant registered in GeneBank
动物细胞中,对铁发生反应的铁蛋白调控主要发生在翻译水平上[48],而植物铁蛋白基因的调控多数发生在转录水平(或转录后水平)[47]。Wei和Elizabeth[48]通过启动子删除,发现在大豆叶中铁蛋白启动子上游序列-304~-218 bp的86 bp是非常必需的,称之为铁调控元件(iron regulatory element,IRE),控制由铁介导的铁蛋白基因去阻遏作用。且IRE和其他任何已知启动子或者IRE之间没有序列同源性,对环境刺激显示出很强的专一性,只有铁离子对铁蛋白表达具有影响,盐胁迫、干旱和H2O2都不起作用。Wicks和Entsch[49]在二倍体豇豆的叶片中发现了3个编码铁蛋白的基因,每个基因都有特定的氨基酸序列并编码一个蛋白质亚基。关于铁蛋白的表达调控问题,已有研究表明,铁在转录和翻译水平上直接调节着铁蛋白的合成[50]。植物中部分调节作用是通过传导途径来实现的,例如,在玉米中,这种调控有2条不同的途径:一是植物激素ABA参与控制基因的表达,二是通过铁和还原信号来调控,在低铁浓度下并有氧化剂如H2O2的参与,并受抗氧化剂的抑制[51,52]。
植物铁蛋白在细胞内由一个小的基因家族编码合成[53]。Jean-Michel等[16]报道拟南芥的4种铁蛋白基因,其中AtFer1和AtFer3的mRNA含量在铁诱导下迅速增加,而AtFer2对铁诱导无任何反应,是由于在AtFer1的5′-UTR区有15 bp的cis调控元件。AtFer2是种子中仅有的一种铁蛋白,其他3种在营养器官中表达,而在种子中未见表达。Fobis等[18]克隆出了玉米铁蛋白的基因ZmFer1和ZmFer2,2个基因均由8个外显子和7个内含子组成,且位置相同,但两者3′端非翻译区和转录起始区及启动子序列有差异,而且也分别受不同的途径调控。铁诱导铁蛋白mRNA的积累是短暂的,在叶中较多,而在根中受铁的影响,则较叶片小。其他一些因素如外源激素和环境胁迫等也诱导铁蛋白基因的表达。而Goto等[54]认为,由于铁蛋白基因是以基因家族的形式存在,则可能存在一类独特的铁蛋白,此种蛋白除了能够结合铁以外,还可以较为容易地结合其他重金属元素。用转基因技术将铁蛋白基因转入植物体内,可以培育出能耐重金属胁迫的植株,这为利用转铁蛋白基因植物净化土壤和水,从而整治环境污染提供了可能。
3.3 植物铁蛋白基因的遗传转化
Goto等[54-56]通过农杆菌介导,将大豆铁蛋白cDNA分别转入水稻、莴苣(Lactuca sativa)、烟草中发现:转入水稻后,在水稻特异性种子存储蛋白谷蛋白启动子GluB-1控制下,只在谷粒中富集铁,其他部位中的铁含量则与正常植株没多大差异,且发现含铁量是对照的1.2~1.7倍。据此,他们认为,这与GluB-1启动子驱动铁蛋白的表达模式有关。并估计这可以满足30%~50%成年人每天对铁的需求;由35S启动子驱动转入莴苣后,转基因植物的生长速度也比野生型的快许多,其鲜重比对照高出27%~42%,且发现铁含量明显提高;而通过CaMV35S启动子将大豆铁蛋白基因转入烟草,其营养组织中铁含量提高30%~40%。
为了提高水稻种子中的铁含量和增强铁的生物利用效率,Lucca等[57]也将菜豆铁蛋白基因转入水稻,同时将曲霉菌(Aspergillus f umigatus)抗热植酸酶引入水稻胚乳,且使富含半胱氨酸的内源金属硫蛋白类似蛋白(metallothionein-like protein)过量表达,发现水稻种子中铁含量提高了2倍,铁吸收的抑制剂植酸也很容易被降解,并且过量的半胱氨酸还能迅速结合铁离子,提高铁的吸收。Georgia等[58]将从大豆中分离的cDNA序列导入水稻和小麦后,再用玉米组成型启动子泛素-1表达,电感耦合氩等离子体光谱法(inductively coupled argon plasma spectrometry,ICAP)分析表明,只是在转基因植物的营养组织中铁水平有提高。水稻叶中比小麦的高一些,其中转基因小麦叶中铁蛋白含量提高50%,而转基因水稻叶中则可以高达2倍。但是在2种植物的种子中都和野生型植株中的铁含量相差不大。徐晓晖等[19]将豌豆铁蛋白基因转入到水稻中,发现转基因水稻的种子和叶片组织中铁含量比非转基因株系的铁含量均有所提高,其中部分株系种子中铁含量比未转基因株系提高了2.3倍。
Van wuytswinkel等[50]用CaMV35S启动子将大豆铁蛋白基因导入烟草后,发现转基因烟草中铁蛋白过量表达,根中铁还原酶活性提高2~5倍,叶片中铁含量增加2~3倍。同时伴随铁吸收的提高,植物对铁的储存能力也相应增加,对由除草剂百草枯引起的氧化胁迫的抗性也增强[31]。Deak等[17]将紫花苜蓿铁蛋白基因导入烟草,获得的转基因烟草具有抵抗由氧化剂如百草枯等引起的氧化胁迫作用的能力,同时还具有抗病毒如烟草坏死病毒(tobacco necrosis virus)、三出交链孢(Alternaria alteoate)、灰葡萄孢(Botrytis cinerea)的抗性。转基因烟草植株由于叶片中积累了铁蛋白,在过量铁或百草枯处理引起的自由基毒害时,还能维持光合作用功能,并对病毒引起的坏死和真菌的感染表现出耐性,蛋白质通过截获细胞中铁,而阻止铁参与Fenton反应,从而保护细胞免受因环境胁迫导致的细胞氧化性损伤。
蔬菜也是人类重要的食品,提高蔬菜中的铁含量对防御铁营养缺乏症和提高小麦、水稻籽实的含铁量一样具有重要意义。菠菜(Spinaciaoleracea)以含铁量丰富而备受欢迎,但其草酸的含量非常高,草酸和钙结合可以引起肾结石,抑制铁的吸收[59],所以发现一种新的含铁量丰富的蔬菜十分必要。2002年,日本科学家成功地将大豆的铁蛋白基因转入莴苣(又称生菜)细胞中,培育开发了可预防贫血症的转基因生菜,其铁含量比一般生菜品种高出约1倍,达370~380 μg/g,这种转基因生菜增强了叶子贮存铁分子的能力,有利于吸收铁元素[60]。
随着近年来的研究,植物铁蛋白转基因研究取得了一定的进展,Beard等[61]于1996年用铁结合蛋白和大豆等食物治疗鼠的铁缺乏症,结果认为使用孟德尔和生物技术途径提高大豆和其他种子中铁结合蛋白的表达和种子铁的其他可溶性成分将有助于全球性缺铁问题的有效解决。Murray Kolb等[62]证明用铁蛋白转基因的水稻饲养小鼠产生的功效与用FeSO4喂养具有相同的功效,两者都可防治贫血,说明采用生物技术实现外源铁蛋白基因在植物中表达,提高自然种子中铁结合蛋白的含量,是一条可持续解决或缓解全球铁营养缺乏的途径。
4 结语
植物中铁蛋白是植物贮存铁的主要形式,而且是生物体获取铁的主要来源,而提高生物体中的含铁量,对加强铁的补给及维持铁平衡有重要意义。铁蛋白作为贮存铁原子、调控铁生物功能、维持铁平衡以及作为一种重要的胁迫反应蛋白,具有抗氧化功能,在植物发育中起重要作用。
目前,对于铁蛋白在植物生理中所起的作用仍然知之不多,很多有关铁蛋白在植物发育过程中的作用关系仍有待于深入研究。对植物不同发育阶段铁蛋白家族的各类成员进行表达分析,是研究和确定这些蛋白在植物生理学中所起作用的必要手段。另外,铁的储存机制和利用铁蛋白基因表达清除氧化胁迫的机制,仍然不十分清楚。铁蛋白结合铁的操纵机制也许能提供一种阻止氧化伤害,增强植物对胁迫耐受性,减轻病害引起损失的新方法。值得引起注意的是:外源转铁蛋白和植物体内源铁蛋白是否可以形成异源多聚体,这种异源多聚体是否具有功能,尚需进一步研究[57]。
抗逆基因工程主要集中在逆境条件下表达的某些基因和抗逆代谢过程中某些酶的研究,其分离、克隆和转化一直是国内外科学家研究的热点,植物抗逆性是十分复杂的性状,一般由多基因调控[63,64]。逆境会引起植物基因表达发生改变,关闭一些正常表达的基因,启动一些与逆境相适应的基因,目前,对某些逆境诱导蛋白的生化功能缺乏了解[65]。因此,植物铁蛋白作为一种胁迫反应蛋白,需要更深入的研究其逆境条件下的生理生化特性,并期待可以通过最新的基因工程、作物育种技术,培育新的抗性作物等,进一步证实铁蛋白生理功能及其关系,加强对植物铁蛋白的研究、应用及推广。
[1]Yona C,Philip B.Iron nutrition of plants in calcareous soils[J].Advances in Agronomy,1982,135:217-240.
[2]Anderson W B.Diagnosis and correction of Fe deficiency in field crops:An overview[J].Journal of Plant Nutrition,1982,5:785-795.
[3]苏振渝.毛皮动物缺铁性贫血[J].北方牧业,2003,24:27.
[4]蔡秋艺,郭长虹,毛文艳,等.植物铁蛋白在人类铁营养中的作用[J].中国农学通报,2007,23:125-128.
[5]Andrews S C,Arosio P,Bottke W,et al.Structure,function and evolution of ferritins[J].Journal of Inorganic Biochemistry,1992,47:161-174.
[6]Harrison P M,Arosio P.The ferritins:Molecular properties,iron storage function and cellular regulation[J].Biochimica et Biophysica Acta,1996,1275:161-203.
[7]Goto F,Yoshihara T,Masuda T,etal.Genetic improvement of iron content and stress adaptationin plants using ferritin gene[J].Biotechnology&Genetic Engineering Reviews,2001,18(3):51-71.
[8]LaulhereJ P,Briat J F.Iron release and uptake by plant ferritin:Effects of pH,reduction and chelation[J].Biochemical Journal,1993,290:693-696.
[9]Briat J F.Metal-iron-mediated oxidative stress and its control[A].In:Montagu M,Inze D,Oxidative Stress in Plants[M].London:Talor and Francis Publishers,2002:171-189.
[10]习阳,刘祥林,黄勤妮.植物铁蛋白转基因的应用[J].植物生理学通讯,2003,39(3):284-288.
[11]Briat J F.Roles of ferritin in plants[J].Journal of Plant Nutrition,1996,19(8-9):1331-1342.
[12]Proudhon D,Briat J F,Lescure A M.Iron induction of ferritin synthesis in soybean cell suspensions[J].Plant Physiology,1989,90:586-590.
[13]Hyde B B,Hodge A J,Kahn A,et al.Studies of phytoferritin identification and localization[J].Journal of Ultrastructure Research,1963,9:248-258.
[14]Lobreaux S,Yewdall S J,Briat J F,et al.Amino-acid sequence and predicted three-dimensional structure of pea seed(Pisum sativum)ferritin[J].Biochemical Journal,1992,288:931-939.
[15]Lescure A M,Proudhon D,Pesey H,etal.Ferritin gene transcription is regulated by iron in soybean cell cultures[J].Proceedings of the National Academy of Sciences,USA,1991,88:8222-8226.
[16]Jean-Michel P,Van Wuytswinkel O,Briat J F,et al.Characterization of an iron-dependent regulatory sequence involved in the transcriptional control of AtFer1 and ZmFer1 plant ferritin genes by iron[J].The Journal of Biological Chemistry,2001,276(8):5584-5590.
[17]Deak M,Horvath G V,Davletova S.Plants ectopically expressing the iron-binding protein,ferritin,are tolerant to oxidative damage and pathogens[J].Nature Biotechnology,1999,17(2):192-196.
[18]Fobis-Loisy I,Loridon K,Lobreaux S,et al.Structure and differential expression of two maize ferritin genes in response to iron and abscisic acid[J].European Journal of Biochemistry,1995,232(3):609-619.
[19]徐晓晖,郭泽建,程志强,等.铁蛋白基因的水稻转化及其功能初步分析[J].浙江大学学报(农业与生命科学版),2003,29(1):49-54.
[20]叶霞.苹果、梨铁蛋白基因的克隆及菜豆铁蛋白基因在转基因苹果和番茄植株中的表达特性研究[D].南京:南京农业大学,2006.
[21]杜仁杰,曲跃军,吴丽丽,等.过量表达铁蛋白基因的转基因烟草抗Co2+能力分析[J].黑龙江医药,2009,22(3):302-306.
[22]Harrison P M,Artymiuk P J,Ford G C,et al.Probing structure-function relations in ferritin and bacterioferritin[J].Advances in Inorganic Chemistry,1991,36:449-486.
[23]Harrison P M,Ford G C,Smith J M A,et al.The location of exon boundaries in the multimeric iron-storage protein ferritin[J].Biology Metals,1991,4:95-99.
[24]Ferreira F,Bucchini D,Martin M E,et al.Early embryonic lethality of H ferritin gene deletion in mice[J].The Journal of Biological Chemistry,2000,275(30):21-24.
[25]Lawson D M,Artymiuk P J,Yewdall S J,et al.Solving the structure of human H ferritin by genetically engineering intermolecular crystal contacts[J].Nature,1991,349:541-544.
[26]Ragland M,Theil E C.Ferritin(mRNA,protein)and iron concentrations during soybean nodule development[J].Plant M olecular Biology,1993,21:555-560.
[27]叶霞,黄晓德,陶建敏,等.转基因苹果组培苗铁蛋白基因在转录水平上的表达[J].果树学报,2006,23(4):491-494.
[28]Roswitha B,Renate M,Dieter N,etal.Excessive iron accumulation in the pea mutants dgl and brz:Subcellular localization of iron and ferritin[J].Planta,1998,207:217-223.
[29]Manuel A M,Lisa M B,Pedro R,et al.Stress-induced legume root nodule senescence:Physiological,biochemical,and structural alterations[J].Plant Physiology,1999,121(1):97-112.
[30]宁春红,杨东鹤,蔡秋艺,等.植物铁蛋白与植物发育[J].黑龙江农业科学,2007,4:103-104.
[31]Seckbach J.Studies on the deposition of plant ferritin as influenced by iron supply to iron-deficient beans[J].Journal of Ultrastructure Research,1968,22:413-423.
[32]Lobr éaux S,Briat J F.Ferritin accumulation and degradation in different organs of pea(Pisum sativum)during development[J].Biochemical Journal,1991,274:601-606.
[33]Huang J S,Barker K R.Influence of Heterodera glucines on leghemoglobins of soybean nodules[J].Phytopathology,1983,73:1002-1004.
[34]Becana M,Moran J F,Iturbe-Ormaetxe I.Iron-dependent oxygen free radical generation in plants subjected to environmental stress:Toxicity and antioxidant protection[J].Plant and Soil,1998,201:137-147.
[35]Balla G,Jacob H S,Balla J,et al.Ferritin:A cytoprotective antioxidant stratagem of endothelium[J].The Journal of Biological Chemistry,1992,267:18148-18153.
[36]Marinos N G.Multifunctional plastics in the meristematic region of potato tuber buds[J].Journal of Ultrastructure Research,1967,17:91-113.
[37]Theil E C,Hase T.Plant and microbial ferritin,and cellular function in animals,plants and microorganisms[J].Annual Review of Biochemistry,1993,56:289-315.
[38]吴平,印莉萍,张立平.植物营养分子生理学[M].北京:科学技术出版社,2001.
[39]刘宝娟,张文兵.铁蛋白的结构、功能及表达调控[J].饲料工业,2009,30(4):42-47.
[40]Alison J W,Ronald E W,Barrie E.Occurrence and expression of members of the ferritin gene family in cowpeas[J].Biochemical Journal,1999,337:523-530.
[41]Sczekan S R,Joshi J G.Isolation and characterization of ferritin from soypeas[J].Journal of Biological Chemistry,1987,262(28):13780-13788.
[42]Vicky B W,Charles A.Leaf senescence in Brassicanapus:Cloning of senescence related genesby subtractive hybridization[J].Plant Molecullar Biology,1997,33(5):821-834.
[43]Jiang T B.Isolation and expression pattern analysis of two ferritin genes in tobacco[J].Journal of Integrative Plant Biology,2005,47:44-47.
[44]郭长虹,王永斌,李丽娅,等.黄花苜蓿铁蛋白基因cDNA的克隆与序列分析[J].哈尔滨工业大学学报,2009,41(3):141-145.
[45]周志钦,成明昊,周泽扬,等.苹果铁结合蛋白基因的克隆和序列分析[J].生物工程学报,2001,17(3):342-344.
[46]Van Wuytswinkel O,Briat J F.Conformational change and in vitro core-formation modifications induced by site-directed mutagenesis of the specific N-terminus of pea seed ferritin[J].Biochemical Journal,1995,305:959-965.
[47]Gaymard F,Boucherez J,Briat J F.Characterization of a ferritin mRNA from Arabidopsisthaliana accumulated in response to iron through an oxidative pathway in dependent of abscisic acid[J].Biochemical Journal,1996,318:67-73.
[48]Wei J Z,Elizabeth C T.Identification and characterization of the iron regulatory element in the ferritin gene of a plant(soybean)[J].The Journal of Biological Chemistry,2000,275(23):17488-17493.
[49]Wicks R E,Entsch B.Functional genes found for three different plant ferritin subunits in the legume,Vigna unguiculata[J].Biochemical and Biophysical Research Communications,1993,192(2):813-819.
[50]Van Wuytswinkel O,Vansuyt G,Grignon N,et al.Iron homeostasisalteration in transgenic tobacco overexpressing ferritin[J].The Plant Journal,1999,17(1):93-97.
[51]Briat J F,Lobreaux S.Iron transport and storage in plants[J].Trends in Plant Science,1997,2:187-193.
[52]Lobreaux S,Thoiron S,Briat J F.Indruction of ferritin synthesis in maize leaves by an iron-mediated oxidative stress[J].The Plant Journal,1995,8:443-449.
[53]Petit J M,Briat J F,Lobreaux S.Structure and differential expression of the four members of the Arabidopsisthaliana ferritin gene family[J].Biochemical Journal,2001,359:575-582.
[54]Goto F,Yoshihara T,Saiki H.Iron accumulation and enhanced growth in transgenic lettuce plants expressing the iron-binding protein ferritin[J].Theoretical and Applied Genetics,2000,100:658-664.
[55]Goto F,Yoshihara T,Shigemoto N.Iron fortification of rice seed by the soybean ferritin gene[J].Nature Biotechnology,1999,17:282-286.
[56]Goto F,Yoshihara T,Saiki H.Iron accumulation in tobacco plants expressing soybean ferritin gene[J].Transgenic Research,1998,7:173-180.
[57]Lucca P,Hurrell R,Potrykus I.Fighting iron deficiency anemia with Iron-rich rice[J].Journal of the American College of Nutrition,2002,21(3):184-190.
[58]Georgia D,Paul C,Eva S.Constitutive expression of soybean ferritin cDNA in transgenic wheat and rice results in increased iron levels in vegetative tissues but not in seeds[J].Transgenic Research,2000,9:445-452.
[59]Theil E C,Burton J W,Beard J L.A sustainable solution for dietary iron deficiency through plant biotechnology and breeding to increase seed ferritin control[J].European Journal of Clinical Nutrition,1997,51(Suppl 4):28-31.
[60]刘燕.红小豆铁蛋白基因的克隆及在水稻中的转化[D].北京:中国农业大学,2005.
[61]Beard J L,Burton J W,Theil E C.Purified ferritin and soybean meal can be sources of iron for treating iron deficiency in rats[J].Journal of Nutrition,1996,126(1):154-160.
[62]Murray Kolb L E,Takaiwa F,Goto F,et al.Transgenic rice is a source of iron for iron-depleted rats[J].Journal of Nutrition,2002,132(5):957-960.
[63]梁哲,姜三杰,未丽,等.三叶草基因工程研究进展[J].草业学报,2009,18(2):205-211.
[64]张振霞,杨中艺,储成才.禾本科牧草与草坪草转基因研究进展[J].草业学报,2004,13(6):92-98.
[65]卢少云,郭振飞.草坪草逆境生理研究进展[J].草业学报,2003,12(4):7-13.
