白桂木生殖物候和繁育系统研究
2010-02-26范繁荣
范繁荣
(福建三明林业学校,福建 三明 365001)
繁殖作为植物有机体生活周期中最为关键、并相对脆弱的一个特殊生活史环节,对生殖成功率和种群的有效更新起着关键性作用,而对植物的生殖物候、花的结构、繁育系统的研究是认识植物生活史的前提[1]。为找出白桂木 (Artocarpus hypargyreus Hance)种群濒危在其生殖物候和繁育系统方面的原因机制,通过对白桂木天然林的定位观测,分析白桂木的生殖物候,在此基础上分别从生殖发育过程、花序结构和繁育系统方面对白桂木有性生殖过程中的濒危环节进行系统研究,分析濒危植物白桂木有性生殖过程的生活史环节,从中发现导致白桂木濒危的某些因素,为揭示白桂木种群的濒危机制和该种群的保护提供科学依据。
1 试验地概况
试验地设在福建省三明市梅列区陈大白桂木天然林内,位于北纬 26°14′~ 26°25′,东经 117°30′~117°47′,属武夷山脉与戴云山脉之间的低山丘陵。为中亚热带季风气候区,有明显的山地气候特征。年平均气温 19.5℃,极端最高气温 42.6℃,极端最低温度 -6.7℃,年均无霜期 272 d~328 d,年均降雨量1 725mm,全年平均日照时数 1 840 h,年平均相对湿度 79%。林地主要成土母岩为花岗岩,土层厚度 1 m以上,土壤为山地红壤。地带性植被为亚热带常绿阔叶林,主要分布树种有甜槠(Castanopsis eyrei)、拟赤杨 (Alniphyllum fortunei)、石栎 (Lithocarpus glabra)、木荷 (Schima superba)、南酸栆 (Choerospondiasaxillaris)等。
2 研究方法
2.1 白桂木生殖物候观测
针对目前国内缺乏白桂木生殖物候资料的现实,本研究参照《中国动植物物候观测年报》(1966-1972,第 3号)的研究方法[2],以福建省三明市梅列区陈大白桂木天然种群为观测对象,进行了为期两年的白桂木生殖物候观测。通过对白桂木天然林的全面踏查,选择生长健壮、并具代表性的白桂木进行定位定株观测。观测时间从 2005年 1月开始至2006年 12月结束,每隔半个月观测 1次(即每月 11日和 26日进行定期观测)。在开花结实期(2006年5月 1日至 2006年 9月 20日)则每两天观测 1次。观察项目有白桂木的形态特征,枝、叶、芽生长的起始时间,花序形态、花芽膨胀、开花、盛花、末花起始时间,果实成熟及起始脱落时间等。
2.2 白桂木杂交指数(Outcrossing index,OCI)的估算
按照 Dafni的标准进行花序直径、花朵大小及开花行为的测量,并进行繁育系统评判[3]。具体方法是:(1)花朵或花序直径< 1mm记为 0;1 mm~2 mm记为 1;2 mm~6mm记为 2;> 6 mm记为 3。(2)花药开裂时间与柱头可授期之间的时间间隔,雌蕊先熟记为 0;雄蕊先熟记为 1。(3)柱头与花药的空间位置,同一高度记为 0;空间分离记为 1。三者之和为 OCI值。评判标准为:OCI=0时,繁育系统为闭花受精(Cleistogamy);OCI=1时,繁育系统为专性自交(Obligate autogamy);OCI=2时,繁育系统为兼性自交(Facultative autogamy);OCI=3时,繁育系统为自交亲和,有时需要传粉者;OCI=4时,繁育系统为部分自交亲和、异交、需要传粉者。
2.3 白桂木花粉 -胚珠比(Pollen-ovule ratio,P/O)的估算
依据 Cruden的标准[4],P/O为 2.7时 ~5.4时,其繁育系统为闭花受精;P/O为 18.1时 ~39.0时,繁弃育系统为专性自交;P/O为 31.9时 ~396.0时,繁育系统为兼性自交;P/O比为 244.7时 ~2 588.0时,繁育系统为兼性异交;P/O比为2 108.0时 ~195 525.0时,繁育系统为专性异交。采用宋志平等方法[82]进行花粉量与胚珠比(P/O)测定。
2.4 白桂木风媒传粉或虫媒传粉的确定或排除
采用重力玻片法进行风媒传粉或虫媒传粉的确定或排除[5]。即以盛花期的植株为花粉源,用涂布凡士林的玻片收集花粉,自花粉源起布片,在树冠中层东西南北四个方向布片,每个方向布 5个玻片,间距 0.5 m,24 h后收片,用 16*10的显微镜进行镜检,取 3次结果的平均值。若镜检结果没发现花粉,则排除风媒传粉的可能。
3 结果与分析
3.1 白桂木开花的物候规律
两年的观测结果表明:白桂木为雌雄同株单性花,雄花的花芽膨胀期开始于 4月下旬,雌花则开始于 5月上旬,止于 6月下旬,其间出现两次现蕾高峰,第 1次在 5月 10日左右,第 2次在 5月 20日左右。从表 1可看出,白桂木的盛花时间为 15 d左右,但其花期长达 3个月左右。观测中发现在当年7月白桂木树上还有零星的雌花序在开放,但此时开放的花发育不良,多不结果,可见开花盛期是白桂木最有效的有性生殖时间。
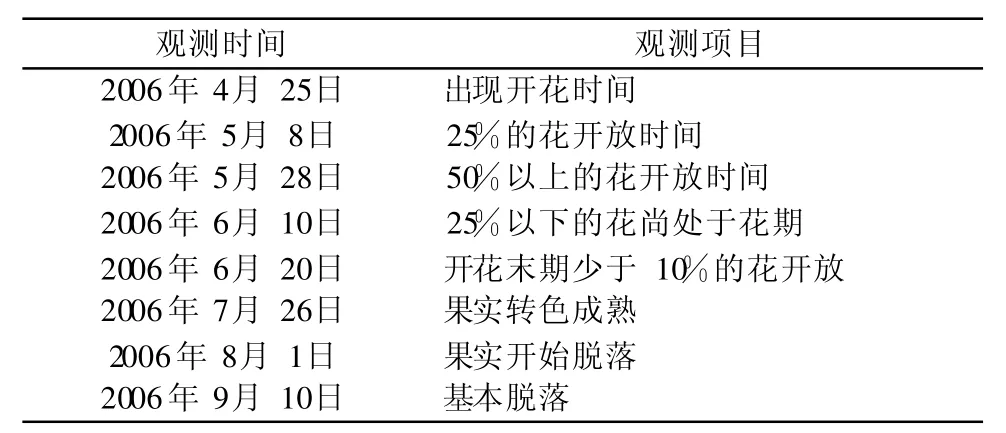
表 1 白桂木的开花结果进程Table 1 Flowering course of Artocarpus hypargyreus
观测中发现:白桂木在盛花期偶见凤蝶停留片刻,但没有发现其它传粉昆虫,而且在试验所布玻片的镜检中均发现有白桂木花粉,因此认为白桂木是存在风媒传粉现象的自花传粉植物。
根据白桂木的开花规律,白桂木属于同步发生花期(Synchonous flowering)植物。所谓同步发生花期是指具有较短的花期但拥有大量的花[6]。Bawa等的研究发现:在植物群落内个体分布较少的物种常通过同步发生花期吸引大量的传粉者,以保证生殖的顺利进行;在群落中乔木层树种以及呈镶嵌格局分布的物种一般也是同步发生花期,风媒植物的花期也常属于同步发生花期[7]。繁育系统通常的定义是代表直接影响后代遗传组成的所有有性特征[8],主要包括花综合特征、花各性器官的寿命、花开放式样、自交亲和程度和交配系统,它们结合传粉者和传粉行为是影响生殖后代遗传组成和适合度的主要因素[9]。观测中发现白桂木在群落中的个体分布较少,为保证生殖的顺利进行,在长期的进化过程中以同时大量开花并进行自花授粉形式来保障生殖成功。
3.2 白桂木的杂交指数与花粉胚珠比
按照 Dafni的方法对白桂木进行杂交指数的测定结果见表 2。白桂木为单性花,同一植株上具有雄花序和雌花序,而且花药散粉初期雌蕊尚未成熟。因此可认为其雄、雌器官在时间和空间上是分离的,其杂交指数(OCI)大于 4,根据 Dafni的标准其繁育系统可以定为异交、部分自交亲和、需要传粉者。

表 2 白桂木的杂交指数Table 2 Outcrossing index(OCI)of Artocarpus hypargyreus
按照宋志平等的方法测定白桂木的花粉 -胚珠比,其结果见表 3。从表 3中可看出:白桂木的 P/O约为 1 248,按照 Cruden的标准,该物种的繁育系统属于兼性异交类型。

表 3 白桂木的花粉胚珠比Table 3 Pollen-ovlue ration of Artocarpus hypargyreus
3.3 白桂木花序的解剖结构
分别将白桂木雄、雌花序进行切片,在电子显微镜下进行观察和拍片,结果见图 1。从图 1中可明显看出:白桂木花序为头状花序,雄花序中单个小花分离,密集着生于球状的花序轴上,雌花序中单个小花相连着生于花序轴上,柱头细小,且周围苞片较多,不易接受花。

图 1 白桂木雄雌花序的切片(×80)Fig.1 Male and female inflorescence's slice(×80)
4 小结
白桂木属于同步发生花期植物,花粉以风媒传播。根据 Dafni的标准所得出的杂交指数(OCI)大于 4,表明白桂木的繁育系统为异交、部分自交亲和、需要传粉者。按照 Cruden的花粉 -胚珠比(P/O)的标准划分,白桂木的花粉 -胚珠比(P/O)约为1248,其繁育系统属于兼性异交类型。
由于白桂木在群落中的个体分布较少,为保证生殖的顺利进行,在长期的进化过程中形成了以同时大量开花的形式来保障生殖成功。但由于开花时期雨水较多,影响花药散粉,而且从花序的解剖结构看,其柱头细小,且受周围苞片的影响,接受花粉量少,同时,由于个体数量少,自交和近交现象严重,这可能是导致白桂木结实率低、种子质量差的原因之一。
[1] 郑宏春,路安民,胡正海.数珠珊瑚(商陆科)的花器官发生(英文)[J].西北植物学报,2004,24(3):476~483.
[2] 中国科学院地理研究所.中国动植物物候观测年报[M].北京:科学出版社,1977:78.
[3] Dafni A.Pollination Ecology[M].Oxford University Press,New York,1992:1~57.
[4] Cruden RW.Pollen-ovule ratio:a conservative indicator ofbreeding systems in flowering plants[J].Evolution,1997,35:1~6.
[5] 宋志平,郭友好,黄双.黄花蔺的繁育系统研究[J].植物分类学报,2000,38(1):53~63.
[6] Gentry AH.Flowering phenology and diversity in tropical Bignoniaceae[J].Biotropica,1974,6:64~68.
[7] Bawa K S,Webb C J.Floral variation and sexual differentiation in Muntingia calabura(Elaeocarpaceae),a species with hermaphrodite flowers[J].Evolution,1983,37(6):1271~1282.
[8] Wyatt R.Pollinator-plantinteractionsand the evolution of breeding systems.In:Real,Led.Pollination biology[M].Orlando:Academy Press,1983,51~59.
[9] 何亚平,刘建全.植物繁育系统研究的最新进展和评述.植物生态学报,2003,27(2):51~163.
