扬子板块伊迪卡拉 (震旦)纪多细胞生物的发展与烃源岩的形成
2010-01-11雷灵芳陈洪德候明才
王 约,雷灵芳,陈洪德,候明才
(1.贵州大学资源与环境工程学院,贵州贵阳 550003;2.成都理工大学沉积地质研究院,四川成都 610059)
扬子板块伊迪卡拉 (震旦)纪多细胞生物的发展与烃源岩的形成
王 约1,雷灵芳1,陈洪德2,候明才2
(1.贵州大学资源与环境工程学院,贵州贵阳 550003;2.成都理工大学沉积地质研究院,四川成都 610059)
生物的生产量以及被保存的沉积有机质是烃源岩形成的重要条件。多细胞生物出现并广泛地生活于扬子地区伊迪卡拉纪陡山沱期晚期的浅海中,生物之间相互依赖和共存的多层次化和复杂化生态系统的建立,使陡山沱期成为扬子地区生命演化和发展重要时期。陡山沱期的黑色泥 (页)岩在沉积时为富含水份的粥性沉积物,同时当时海洋中的氧化-还原界面较浅,生物遗体可迅速地被沉积物所覆盖而形成烃源岩。伊迪卡拉纪陡山沱期是化学、气候和环境变化的剧烈时期,也是扬子地区生命演化、有机质生产力发展和烃源岩形成的一个重要时期。
烃源岩;有机质;氧;多细胞生物;陡山沱期;扬子板块
有机质的丰度是烃源岩评价的一个重要方面,而沉积有机质则源于生物及其代谢的产物。因而生物的生产量以及被保存的沉积有机质丰度成为烃源岩形成的重要条件。
“雪球事件”(南沱冰期)之后的伊迪卡拉 (震旦)纪陡山沱期是化学、气候和环境变化的剧烈时期,是多细胞生物起源与发展的关键时期,也是Rodinia超大陆汇聚和裂离的重要地质时期,同时,也是扬子地区的陡山沱期物质沉积方面的重要转折时期。
1 多细胞生物起源和发展
一种古老的丝状的原核细胞化石被发现于南非太古界 Swaziland超群 Onverwacht群和西澳大利亚的 Pilbara超群 Warrawoona群的燧石和碳质燧石中[1~3]。被解释为真核细胞生物标志化合物的甾烷发现于北澳大利亚的 Barney Creek组的沥青中 (约1.7Ga)[4],华北长城系串岭沟组中产较大的球形微化石[5],真核细胞在1.7~1.9Ga前已为生态系中的重要成员[6]。多细胞植物的宏体碳质压膜化石在华北长城系常州沟组 (约1.8Ga)[7]和团子山组 (约1.7Ga)中被发现[8~10]。前伊迪卡拉纪的多细胞植物发现于加拿大[11]、美国[12]、澳大利亚[13]、以及中国[14,15]等地区。尽管分子生物钟方法推算的多细胞动物的最早时间可达1.2Ga[16],但前伊迪卡拉纪多细胞动物的直接证据较为稀少且有所争议[17~19]。
从原核单细胞生物到真核单细胞生物再到多细胞生物的演化与发展,是地球表层生物有机物丰度提高的重要基础。华北中、上元古界从串岭沟组至下马岭组泥 (页)岩中的有机碳平均含量由小于1%到大于3%的变化[20],从一个方面反映了生物的演化发展与有机质丰度的密切相关性。另外,青白口系下马岭组中存在由红藻堆积形成的、有机碳含量为21.41%~22.91%的油页岩[21],表明了多细胞生物对烃源岩形成的贡献。
以多细胞生物为特征的新元古代伊迪卡拉纪生物群被发现于全球的许多地区:澳大利亚[17]、纽芬兰[22]、纳米比亚[23,24]、俄罗斯[25]、英国[26]、乌克兰[27]、加拿大[28]、美国[29]和中国等。伊迪卡拉纪是多细胞生物起源、演化与发展的重要时期,也正是这一时期,在东欧[30]和西北利亚[31]等地区形成了生油的烃源岩。
2 有机质丰度与大气含氧量
众所周知,现生生物绝大多数是依赖氧而进行新陈代谢、生活和繁殖的。太古宙的地球表层大气的含氧量可能相当于现代含氧量水平 (PAL)的1‰以下[32,33];元古宙大气含氧量上升为 1%~10%PAL[33];新元古代的大气含氧量再次上升到20%PAL左右[34]。生物的演化与发展与大气含氧量的变化有较密切的相关性 (图1)。Nursall(1959)[35]曾提出元古宙大气氧分压 (ρO2)的增加使宏体多细胞生物的进化成为可能;一些学者进一步认为元古宙末ρO2高于1%PAL时,使生物需氧新陈代谢成为可能,并使大气圈的臭氧达到具有屏蔽太阳紫外线以保护多细胞生物生活和繁殖的意义[36~38]。同时,环境中具有足够支持微生物呼吸的游离氧是维持宏体多细胞生物生活和繁殖的基础,
而且多细胞生物的繁盛也为微生物提供了丰富的有机营养质来源,促进微生物的繁盛。因而,大气含氧量和海水中游离氧量的升高是地球表层有机质丰度升高的一个重要基础。早古生代多细胞生物的快速发展以及大多数烃源岩层中 (如寒武系的牛蹄塘组,奥陶系的五峰组,志留系的龙马溪组等)均产有丰富的需氧多细胞生物等表明有机质丰度与氧的内在关联。
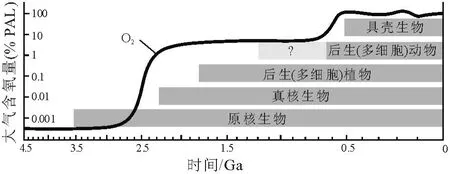
图 1 大气含氧量变化与生物演化 (据 Kump,2008[34]修改)Fig.1 Variationsin atmosphere oxygen and organic evolution(modified from Kump,2008)
3 扬子板块陡山沱期的多细胞生物与有机质的发展
3.1 多细胞生物与氧
“雪球事件”之后的陡山沱期,气候开始转暖,海水温度升高,为生物的发展提供了良好的条件。在华南,先后报道的以球状立体形态保存的多细胞生物[39~42]和以碳质压膜化石方式保存的宏体多细胞生物[42~52],表明陡山沱期的扬子地区广泛地生活有较为丰富的多细胞生物,这些多细胞生物成为改善扬子地区生物生活环境的重要基础和原动力。特别是陡山沱期晚期,生活于扬子海中丰富的宏体多细胞植物多具有营固着作用的固着器或拟根和营光合作用为主的叶状体 (图 2A,2B,2C,2D,2E,2F,2N,2O),表明它们营固着生活方式,并呈半浮状直立于海底。这些半浮状直立生长的宏体藻类,打破了平卧生活方式微生物的统治,大幅度地提高了光合作用的效率,增大了海水中的含氧量[43,44,48,49,53]。特别是分枝的藻类 (图 2E)在陡山沱期晚期的出现和丰富,加大了宏体多细胞植物之间对于阳光的竞争力,同时也增加了氧量的释放。值得一提的是Runnegar(1982)[54]估计伊迪卡拉生物群中的两侧对称动物 Dickinsonia至少需 pO2在6%~10%PAL方能存活,而生活于扬子地区陡山沱期沉积物表面的环节动物 Wenghuiia(图 3A,3B,3C)[51~53]表明底层水体中的含氧量已达到较高的水平。陡山沱期晚期氧量的增加在地球化学研究方面也有相似的结论[55~57],Kump(2008)[34]估计当时的大气含氧量为20%PAL左右。氧量的增加逐渐改善了海洋水体的环境质量,促进了生物的发展,增加了海洋中有机质的丰度。因而陡山沱期成为扬子地区生命演化和发展的重要转换时期。
3.2 多层次、复杂化生态系统与有机质丰度
宏体多细胞植物在陡山沱期的大量出现,也成为多细胞动物新的食物来源,从而改变了动物的食谱,为动物的快速演化和发展提供了一个新的和良好的生活环境和生态链基础[43,49]。
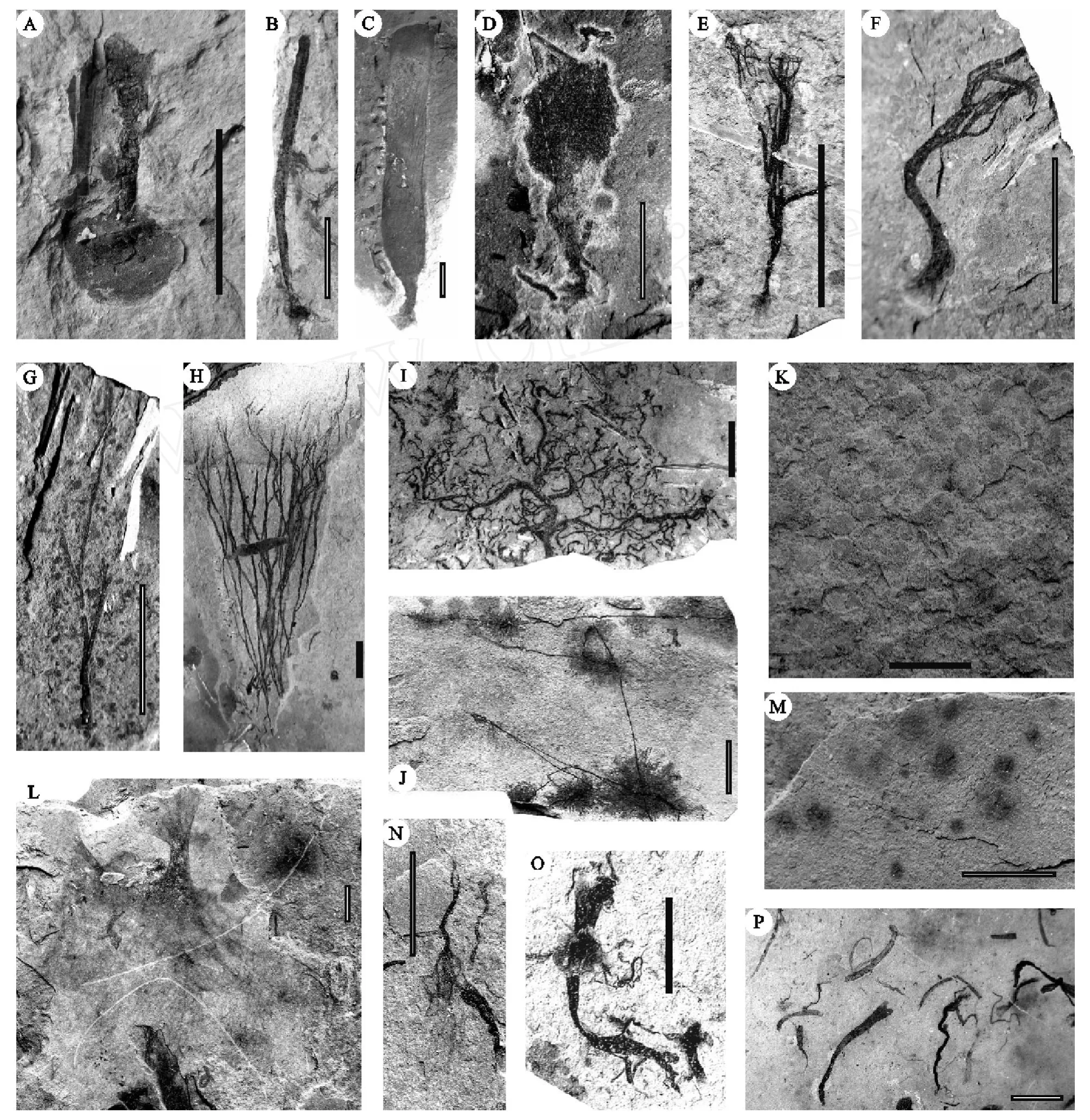
图 2 伊迪卡拉纪陡山沱期的宏体多细胞植物A.具圆盘状固着器的未命名藻类;B.不分枝的的Baculiphyca taeniataYuan et al.,分叶状体和固着器;C.不分枝的藻类 Gesinella hunanensis Steiner et al.,叶状体表面见V字形丝状藻丝的;D.具拟根的扇状藻类 Sectoralga wenghuiensisWang et al.;E,F.具拟根的不等二分枝藻类Zhongbaodaophyton robustusWang et al.;G.不等二分枝藻类 Zhongbaodaophyton crassaChen et al.;H.等二分枝藻类 Doushantuophyton lineare Chen;I.多次不等二分枝的 Jiangkouphyton guizhouensisWang et al.;J.藻丝团与成束状藻丝相连的藻类 Globusphyton lineareWang et al.;K.大小不一而密集的圆币状藻类Beltanellifor m is brunsaeMenner;L.叶状体宽大的未命名藻类;M.圆形或卵圆形的藻类Chuaria circularisWalcott;N.保存良好的丝状拟根;O.呈束状的丝状拟根与叶状体保存在同一层面;P.示宏体藻类具有定向性排列.线段比例尺长10mmFig.2 Macroscopic metaphytes during the Doushantuoan(Ediacaran)A.Unnamed algae;B.Baculiphyca taeniataYuan et al.;C.Gesinella hunanensisSterner et al.;D.Sectoralga wenghuiensisWang et al.;E.&F.Zongbaodaophyton robustusWang et al.;G.Zongbaodaophyton crassaChen et al.;H.Doushantuophyton lineare Chen;I.Jiangkouphyton guizhouensisWang et al.;J.Globusphyton lineareWang et al.;K.Beltanellifor m is brunsaeMenner;L.Unnamed algae;M.Chuaria circularisWalcott;N.Well-preserved filamentous algal roots;O.Filamentous algal roots and thallus on the same bedding plane;P.Oriented arrangement ofmacroscopic algae.Scale:10 mm
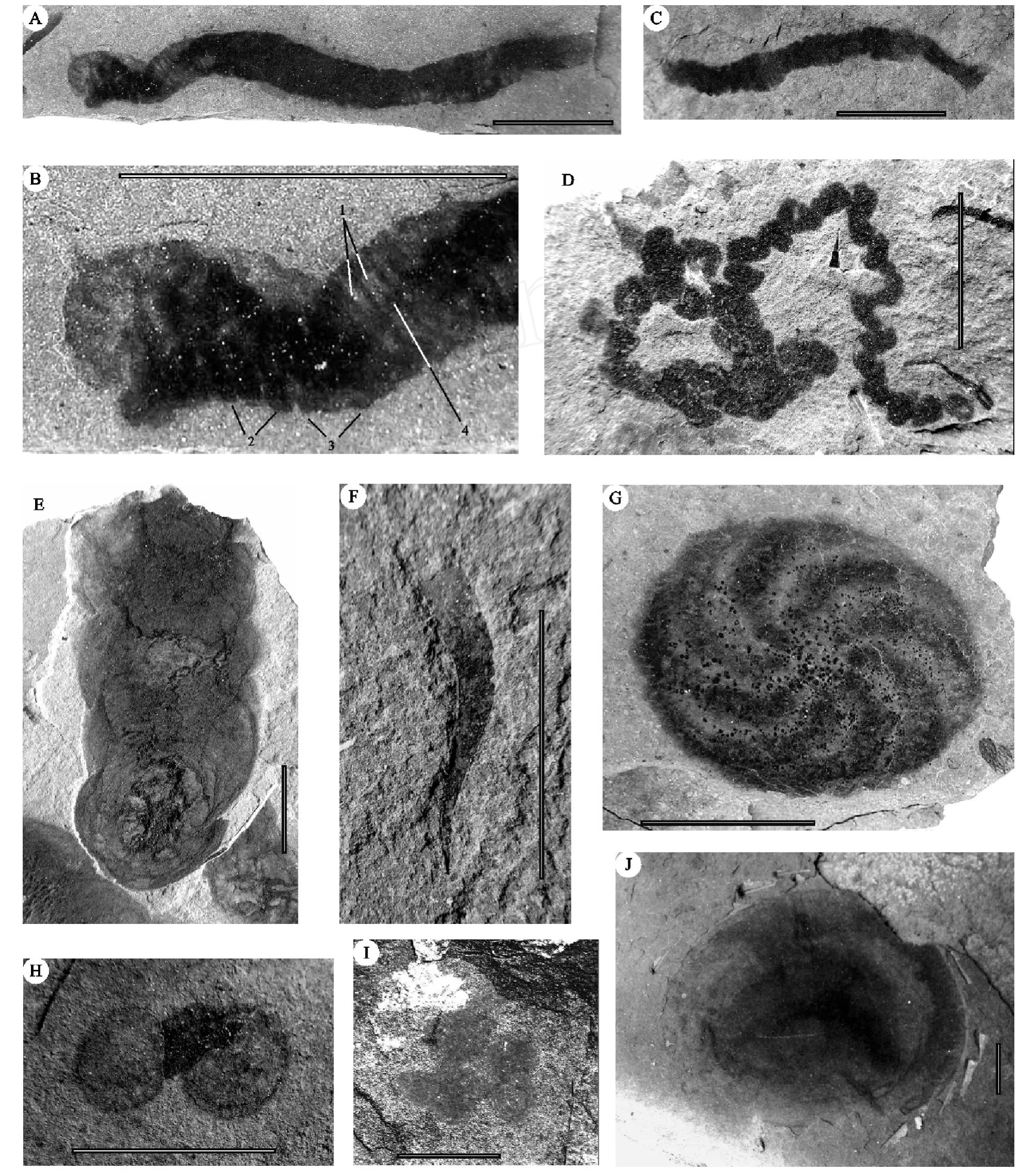
图 3 伊迪卡拉纪陡山沱期的宏体多细胞动物A,B,C.环节动物W enghuiiaWang andWang。B,示图A身体前部,1,体节,2,拟疣足,3,刚毛,4,消化道;D.具珠状构造和边缘乳头状构造的遗迹化石LinbotulichnusLi and Ding;E.具分叉和结网海绵丝的海绵动物 CucullusSteiner;F.两侧对称动物 ProtoconitesChen et al.;G.具栉板和纤毛的八辐射对称栉水母类动物 EoandromedaTang et al.;H.圆盘状具同心纹的未命名水母类动物;I.叶脊三辐射对称的三叶动物Trilobozoa Fedonkin;J.圆盘状具有中央小瘤和同心褶的水母类动物 CyclomedusaSprigg.线段比例尺长10mmFig.3 Macroscopic metazoans during the Doushantuoan(Ediacaran)A,B&C.W enghuiiaWang and Wang;D.LinbotulichnusLi and Ding;E.CucullusSteiner;F.ProtoconitesChen et al.;G.Eoandrom edaTang et al.;H.Unnamed medusae;I.Trilobozoa Fedonkin;J.Cyclom edusaSprigg.Scale:10 mm
在陡山沱期的扬子海中,一些宏体多细胞动物,如囊状的Cucullus(图 3E)和锥状的Protoconites(图 3F),虽未见明显的固着器,但它们可能是以其基部插入沉积物中而固着[42,44,47~49,53],构成了扬子陡山沱期生物群主体面貌,展现出一种“原始海底草原 ”的景观[43,44,48,49]。而紧密成群的圆币状Beltanellifor m is(图2K)和匐葡状生长的Globusphyton(图 2J)等宏体藻类以及可能的平卧微生物生活于沉积物表面,一些盘状底栖的后生动物Cyclom edusa(图3J)和三叶动物 (Trilobozoa)(图 3 I)[25,58,59]等活动于的海床上,另有一些两侧对称的环节动物W enghuiia(图 3A-3C)和遗迹化石Linbotulichnus(图 3D)造迹生物 (可能为蠕形动物)等多细胞动物蜿蜒爬行、觅食于海底的沉积物表面。另外,浮游藻类如宽大叶状体的藻类 (图 2L)和Chuaria(图 2M)及可能的浮游微生物构成的水体上层群落;具有规律排列羽状纤毛和中央构造的盘状Eoandrom eda(图 3G)可能系浮游类型的栉水母类(Ctenophora)[52,53]。
食物链是维系生态系统存在和发展的重要纽带。在陡山沱期的生态系统中,生产者和分解者与前伊迪卡拉纪时相似,拥有丰富的微体和宏体植物,所不同的是,陡山沱期晚期已有叶状体更为宽大和分枝的宏体植物,表现出对阳光较强的竞争态势。同时,华南陡山沱期晚期还有相当数量的具有消化系统的蠕虫动物和环节动物[43,45,51~53]以及造迹的蠕虫类动物[45,47~49],其是一类摄食性的消费者。与主要为化学共生作用生活方式的前伊迪卡拉纪[53,60]生物相比较,摄食性生物可使食物 (供体生物)中的相当部分能量在生物体内有效地传递,建立起较高效能的能量传递营养构造[53]。另外,华南陡山沱期晚期的生态系统包含有分枝宏体植物、浮游多细胞宏体生物和摄食性动物,不仅在生物生活空间和对环境的改造上,在生物间对环境因子需求的相互依存和相互竞争上,而且在食物链和营养构造等方面构成具有多层次化和复杂化的生态金字塔模式[53]。这一生态系统 (图 4)的建立,成为生物演化和发展的一个关键时期,不仅增强了生物之间的相互依赖和共存的能力,也增强了生物群自身抵抗环境变化并连续、快速繁盛和发展的能力,同时也增大了生物 (包括多细胞生物和微生物)的多样性,增加了海洋中有机质的丰度。
4 大地构造演化、沉积物质变化与有机质保存
南华纪正是新元古代 Rodinia超大陆发育的高峰阶段[61~64]。扬子板块的前南华系为变质岩岩系,被称为“似盖层”[65],南华系主要以冰碛砾岩、杂砂岩、砂岩等成熟度不高的碎屑岩为主,有机质的保存条件较差,仅在川东南、渝南、黔东北和湘西北一带有间冰期 (大塘坡期)发育的厚度较小、分布有限的黑色页岩的沉积。“雪球事件”之后的陡山沱期,扬子地区发展成为独立的古陆[62,63];同时,湖南石门陡山沱期的古纬度为3.0±4.5°的近赤道地区[66]。处于赤道低压带内的扬子板块,气温较高、湿度较大、雨量充沛,主要以碳酸盐岩和黑色页岩的沉积为主,特别是黑色和暗色泥 (页)岩、硅质岩以及磷质岩岩系广泛地分布于扬子地区。陡山沱期沉积物质、特别是沉积物质粒度的陡然变化与 Rodinia超大陆的离散密切相关,反映出陡山沱期化学、气候和环境等的剧烈变化。另一方面,这一剧烈的变化为陡山沱期的生命、特别是为多细胞生命的起源和发展提供了良好的水温、丰富的无机营养盐环境等条件,成为陡山沱期烃源岩形成所需的丰富有机质来源的重要基础。再一方面,沉积物质粒度的变化以及立体生态系统所产出的丰富微粒状和胶体状有机质沉积物有利于有机质的保存。

图 4 伊迪卡拉纪陡山沱期的立体生态系统及海洋中的还原性示意图w.水体;d.粥性沉积物;re.还原强度Fig.4 Sketch to show the three-d imensional ecosystems and the reduction in oceans during the Doushantuoan(Ediacaran)w=watermass;d=gruel sediments;re=reduction intensity
5 埋藏方式与烃源岩的形成
黑色或暗色泥 (页)岩系常被视为静水还原环境,甚至被认为与深海及远洋沉积相关。许多学者认为陡山沱晚期宏体生物群生活于滞流缺氧富硫的还原环境[42,43,45,47]。保存于黑色泥 (页)岩中的陡山沱期多细胞宏体化石保存完整,特别是宏体藻类的丝状拟根被完整保存 (图 2D,2E,2N),说明它们是未被长距离搬运的原地或近原地埋藏。但多细胞藻类的丝状拟根呈束状与叶状体呈线状常保存在同一层面 (图 2N,2O)以及定向性排列的标本 (图 2P)等保存特征表明它们又是经过一定的水流作用改造后而被埋藏的。宏体藻类发育的固着器或拟根(图 2A~2F,2N,2O),也反映出具水动力环境的生态特征。现代海洋中藻类植物的生长不仅需要充沛的阳光,也需要丰富的无机营养盐供应[68]。一定能量水动力条件的存在有利于生物释放氧的扩散和无机盐的供给,有利于整个生物群的生活、生长和繁殖。王约和王训练 (1996)[68]通过对宏体藻类化石碎片的定向测量和显微镜下对黑色泥 (页)岩微层理的研究,认为陡山沱期宏体生物的生长具有周期性,并可能与洋流的影响有关。宏体多细胞动物的生活与繁殖需一定的含氧量,特别是生活于沉积物表面的环节动物W enghuiia和遗迹化石Linbotulichnus造迹生物的存在,意味着当时水体底部已有一定的含氧量水平。另外,丁莲芳等(1996)[45]曾依据庙河生物群赋存围岩的 d18O判别其水介质条件为正常盐度的海水。因而陡山沱期宏体多细胞生物生活于一片温暖清澈、正常盐度、光照充分、具一定水动力条件、富有无机营养盐和有一定含氧量水平的相对平静浅海环境。将黑色或暗色泥(页)岩简单地视为某一还原静水环境或与深海和远洋沉积相关可能欠妥。一个完全还原和静水的环境通常不宜于生物的大量生长和繁殖。伊迪卡拉纪大气的 CO2的浓度较高,但 O2的含量已达一定的水平[6,33~38]。陡山沱期扬子海中已形成一个多层次和复杂化的生态系统,也表明了水体的氧含量已达到可供生物生活和繁殖所需新陈代谢的氧量。
通过对宏体藻类固着器或拟根及黑色页岩微层理的研究,王约和王训练 (2006)[68]得出这样的结论:在不考虑成岩过程和成岩后变化的情况下,陡山沱期的宏体多细胞生物固着生长的基底为含水分近80%的沉积物,即为泥质沉积物颗粒之间几乎没有接触而充水的粥性基底 (soup-ground)。宏体藻类圆盘状的固着器 (图 2A)也是生物适应这一粥性基底底质环境的生态特征。实际上,泥 (页)岩在沉积时多含有较高的水分,其中的水分作为沉积物的一部分使沉淀的较少颗粒物质可形成较厚的沉积物。随埋藏深度的加大,沉积物和水体界面之下的沉积水分与海水水体的交换量逐渐减小,逐渐形成滞流还原的环境 (图 4)。也就是说,在富含有宏体多细胞生物的陡山沱组黑色页岩沉积时,海洋水体是有氧环境,含氧量从海底至水体近表层逐渐升高,而沉积物 (包括其沉积的水分)内部则是还原环境。
水体中和沉积物表层的生物排泄物以及死亡后的生物遗体沉积于沉积物的表面或被埋藏于沉积物的表层。一方面,尽管海底存在一定量的氧,但陡山沱期大气中的含氧水平相对较低,生物遗体不易被氧化而腐烂分解。另一方面,一定量的颗粒物质(包括生物排泄物和生物遗体)沉淀就可形成较厚的粥性沉积物,可较迅速地将生物遗体完全覆盖,而富含水分的粥性沉积物内部为一还原的环境。再一方面,被沉积物覆盖的生物遗体在腐烂和分解过程中,H2S等物质增强了生物遗体周围的还原性,导致生物遗体处于较强的还原环境中,生物遗体得以良好保存,也使有机物质得以保存而形成黑色或暗色的泥 (页 )岩,即烃源岩。
陡山沱组黑色页岩的有机质类型主要为腐泥型,有机碳含量为 1%~4%,最高可达 15.47%[69]。另外,沥青质砾石被发现于川北青川一带的早寒武世筇竹寺期的碎屑岩地层中[70]。下寒武统的沥青质砾石表明,在沥青砾石沉积之前 (即前寒武纪)川北、青川地区已存在有古油藏,即前寒武纪陡山沱期的地层中可能存在有规模性生烃的烃源岩[70]。
5 结 语
(1)生物物质的生产量以及被保存的有机质丰度是烃源岩形成的重要条件。“雪球事件”之后的伊迪卡拉纪陡山沱期是化学、气候和环境变化的剧烈时期,是多细胞生物起源与发展的关键时期,同时也是陡山沱期扬子地区物质沉积的重要转折时期。
(2)多细胞生物广泛地生活于扬子地区陡山沱期的海洋中,其中半浮状直立生长的宏体藻类大幅度地提高了光合作用的效率,使海水中的含氧量达到维持宏体多细胞生物生活和繁殖的水平。氧量的增加同时也改善了海洋水体的环境质量,促进了生物的发展。一个生物之间相互依赖和共存的多层次化和复杂化生态系统的形成,使陡山沱期成为扬子地区生命演化和发展的关键时期。
(3)黑色页岩和黑色硅质岩以及磷质岩系广泛地分布于陡山沱期的扬子浅海中。受 Rodinia超大陆的离散构造运动影响,一方面陡山沱期沉积物质发生陡然的变化,另一方面陡山沱期生命、特别是为多细胞生命的分异度和丰度迅速提高,成为陡山沱期丰富有机质来源和保存条件的重要基础。
(4)陡山沱期大气中的含氧水平相对较低,同时陡山沱期的黑色泥 (页)岩在沉积时为沉积物颗粒之间几乎没有接触而充水的粥性沉积物。生物死亡后的遗体不易腐烂和分解,而可较迅速地被粥性沉积物所覆盖,导致生物遗体处于较强的还原环境中,使有机物质得以保存而形成烃源岩。
[1] AWRAM IK S M,SCHOPF J W,WALTER M R.Filamentous fossil bacteria from the Archean of Western Australia[J].Precambrian Research,1983,20:357-374.
[2] SCHOPF J W,PACKER B.Early Archean(3.3-billion-3.5-billion-year-old)microfossils from the Warrawoona Group,Australia[J].Science,1987,237:70-73.
[3] SCHOPF JW.The oldest fossils andwhat theymean[A].Schopf J W.Major Events in the History ofLife[C].Boston:Jones and Bartlett,1992.29-63.
[4] SUMMONS R E,POWELL T G,BOREHAM C J.Petroleum geology and geochemistry of the middle Proterozoic McArthur Basin,NorthernAustralia,ⅢCompositionofextractable hydrocarbons[J].Geochimica et Cos mochimica Acta,1988,52:1747-1763.
[5] 阎玉忠,刘志礼.论长城系微化石群的真核生物意义[J].微体古生物学报,1993,10(2):167-180.
[6] KNOLL A H.The early evolution of eukaryotes:a geological perspective[J].Science,1992,256:622-627.
[7] 朱士兴,孙淑芳,黄学光,等.燕山常州沟组 (约 1800Ma)碳质压型化石及其多细胞组织的发现 [J].科学通报,1999,44(14):1552-1557.
[8] 杜汝霖,田立富,李汉棒.蓟县长城系高于庄组宏观生物化石的发现[J].地质学报,1986,60(2):115-120.
[9] ZHU S,CHEN H.Megascopic multicellular organis ms from the 1700-million-old Tuanshanzi Formation in the Jixian area,North China[J].Science,1995,270:620-622.
[10] 阎玉忠,刘志礼.中国蓟县长城系团山子宏观藻群[J].古生物学报,1997,36(1):18-41.
[11] HOFMANN H J.The mid-Proterozoic Little Dal macrobiota,Mackenzie Mountains,northwest Canada[J].Palaeontology,1985,28:331-354.
[12] WALTERM R,OEHLER J H,OEHLER D Z.Megascopic algae 1300 million years old from the Belt Supergroup,Montana:a reinterpretation of Walcott'sHelm inthoidichnites[J]. Journal of Paleontology,1976,50(5):872-881.
[13] GREY K,W ILL IAMS I R.Problematic bedding-plane markings from the Middle Proterozoic Manganese Subgroup,Bangemall Basin,Western Australia[J]. Precambrian Research,1990,46:307-327.
[14] 陈孟莪,郑文武.先伊迪卡拉期和淮南生物群[J].地质科学,1986,7(3):221-231.
[15] 杜汝霖,田立富.燕山地区青白口系宏观藻类化石及其地层意义[J].前寒武纪地质,1986,(3):495-511.
[16] WRAY G,LEV INTON J S,SHAPI RO L H.Molecular evidence of deep Precambrian divergences among the metazoan phyla[J].Science,1996,274:568-573.
[17] GLAESSNER M F.The dawn of animal life,a biohistorical study[M].Cambridge:Cambridge University Press,1984.1-236.
[18] WALTER C D,DU R,HORODYSKIR J.Coiled carbonaceous megafossils from the middle Proterozoic of Jixian(Tianjin)and Montana[J].American Journalof Science,1990,290-A:133-148.
[19] 王约,徐一帆.贵州瓮安埃迪卡拉系陡山沱组上段底部和下段的遗迹化石[J].现代地质,2007,21(3):469-478.
[20] 王杰,陈践发,王大锐,等.华北北部中、上元古界生烃潜力及有机质碳同位素组成特征研究[J].石油勘探与开发,2002,29(13):13-15.
[21] 张水昌,张宝民,边立曾,等.8亿多年前由红藻堆积而成的下马岭组油页岩[J].中国科学D辑,地球科学,2007,37(5):636-643.
[22] ANDERSON M M,M ISRA S B.Fossils foundinthe Precambrian Conception Group of southeastern Newfoundland[J].Nature,1968,220:680-681.
[23] GERMS G J B.Possible new sprigginid worm and a new trace fossil from the Nama Group,Southwest Africa[J].Geology,1973,1:69-70.
[24] JENK INS R J F.The enigmatic Ediacaran(late Precambrian)genus Rangea and related forms[J].Paleobiology,1985,11:336-355.
[25] FEDONK IN M A.Systematic description of Vendian Metazoa[A].Sokolov B S,Ivanovskij A B.The Vendian System:Palaeontology[C].Berlin:Springer-Verlag,1990.71-120.
[26] COPE J W.Precambrian fossil of the Carmarthen area,Dyfed[J].Nature inWales,1983,1:11-16.
[27] RUNNEGAR B.Evolution of the earliest animals[A].Schopf J W.Major Events in the History ofLife[C].Boston:Jones and Bartlett,1992.65-93.
[28] NARBONNE G M,HOFMANN H J.Ediacaran biota of the Wernecke Mountains,Yukon,Canada[J]. Palaeontology,1987,30:647-676.
[29] G IBSON G G,TEETER S A,FEDONK IN M A.Ediacaran fossils from the Carolina slate belt, Stanly County,North Carolina[J].Geology,1984,12:387-390.
[30] BAZHENNOVA O K,AREFIEV O A.Geochemical peculiarities of pre-Cambrain source rocks in the East European Platform[J].Organic Geochimistry,1996,25:341-351.
[31] FOWLER MG,DOUGLAS AG.Saturated hydrocarbon biomarkers in oils ofLate Precambrain age from Eastern Siberia[J].Organic Geochimistry,1987,11:201-203.
[32] KAST ING J F,HOWARD M T.Atmospheric composition and climate on the early Earth[J].Philosophical Transactionsof the Royal SocietyB:Biological Sciences,2006,361:1733-1742.
[33] HOLLAND H D.The oxygenation of the atmosphere and oceans[J].Philosophical Transactions of the Royal Society of London,2006,361:903-915.
[34] KUMP L R.The rise of atmospheric oxygen[J].Nature,2008,451:277.
[35] NURSALL J R.Oxygen as a prerequisite to the origin of the metazoa[J].Nature,1959,183:1170-1172.
[36] BERKNER L V,MARSHALL L C.On the origin and rise of oxygen concentration in the earth's atmosphere[J].Journal of Atmospheric Science,1965,22:225-261.
[37] TOWE K M.Oxygen-collagen priority and the early metazoan fossil record[J]. Proceedings of the National Academy of Science USA,1970,65:781-788.
[38] KAST ING J F,HOLLAND H D,KUMP L R.Atmosheric evolution:the rise of oxygen[A].Schope J W,Klein C.The Proterozoic Biosphere, a Multidisciplinary Study [C].Cambridge:Cambridge University Press,1992.159-163.
[39] L I C,CHEN J,HUA T.Precambrian sponges with cellular structures[J].Science,1998,279:879-882.
[40] XI AO S,ZHANG Y,KNOLL A.Three-dimensionalpreservation of algae and animal embryos in a Neoproterozoic phosphorite[J].Nature,1998,391:553-558.
[41] Y IN L,ZHU M,KNOLL A H et al.Doushantuo embryos preserved inside diapause egg cysts[J].Nature,2007,446:611-663.
[42] 袁训来,肖书海,尹磊明,等.陡山沱期生物群 -早期动物辐射前夕的生命[M].合肥:中国科学技术大学出版社,2002.26-133.
[43] 陈孟莪,萧宗正,袁训来.晚震旦世的特种生物群落-庙河生物群新知[J].古生物学报,1994,33(4):391-403.
[44] 陈孟莪,陈其英,萧宗正.试论宏体植物的早期演化[J].地质科学,2000,35(1):1-15.
[45] 丁莲芳,李勇,胡夏嵩,等.震旦纪庙河生物群 [M].北京:地质出版社,1996.28-33.
[46] TANG F,YI N C Y,BENGTSON S et al.A new discovery of macroscopic fossils from the Ediacaran Doushantuo Formation in the Yangtze Gorges area[J].Chinese Science Bulletin,2006,51(12):1487-1493.
[47] XIAO S,YUAN X,STEINER M et al.Macroscopic carbonaceous compressions in a ter minal Proterozoic shale:A systematic reassessment of the Miaohe biota,South China[J].Journal of Paleontology,2002,76:347-376.
[48] 王约,何明华,喻美艺,等.黔东北震旦纪陡山沱晚期庙河型生物群的生态特征及埋藏环境初探[J].古地理学报,2005,7(3):327-335.
[49] 王约,王训练,黄禹铭.黔东北伊迪卡拉纪陡山沱组的宏体藻类[J].地球科学,2007,32(6):828-844.
[50] 王约,王训练,黄舜铭.华南伊迪卡拉系陡山沱组Protoconites的分类位置与生态初探[J].地球科学,2007,32(Sup.):41-50.
[51] WANG Y,WANG X L.Annelid fromthe Neoproterozoic Doushantuo Formation in the Northeast Guizhou,China[J].Acta Geologica Sinica,2008,82(2):257-265.
[52] WANG Y,WANG X L,HUANG YM.Megascopic symmetrical Metazoans from the Ediacaran Doushantuo Formation in the Northeastren Guizhou,South China[J]. Journal of China University of Geosciences,2008,19(3):200-206.
[53] 王约,赵明胜,杨艳飞,等.华南陡山沱期晚期宏体生物生态系统的出现及其意义[J].古地理学报,2009,11(6):640-650.
[54] RUNNEGAR B.Oxyen requirements,a biology and phylogenetic significance of the late Precambrian wor m Dickinsonia and the evolution of the burrowing habit[J].Alcheringa,1982,6:223-239.
[55] 张同钢,储雪蕾,张启锐,等.陡山沱期古海水的硫和碳同位素变化[J].科学通报,2003,48(8):850-855.
[56] 郭庆军,杨卫东,刘丛强,等.贵州瓮安生物群和磷矿形成的沉积地球化学研究 [J].矿物岩石地球化学通报,2003,22(3):202-208.
[57] 吴凯,马东升,潘家永,等.贵州瓮安磷矿陡山沱组地层元素地球化学特征[J].华东理工学院学报,2006,29(2):108-114.
[58] FEDONK IN M A.Precambrian metazoans:the problems of preservation,systematics and evolution [J]. Philosophical Transactions of the Royal Society ofLondon,1985,311:27-45.
[59] SE ILACHER A.Biomat-related lifestyles in the Precambrian[J].Palaios,1999,14:86-93.
[60] MCCALL G J H.The Vendian(Ediacaran)in the geological record:Enigmas in geology’s prelude to the Cambrian explosion[J].Earth-Science Reviews,2006,77:1-229.
[61] POWELL C MCA,L I Z X,MCELH INNY M Wet al.Paleomagnetic constraints on the Neoproterozoic Breakup of Rodinia and the Mid-Cambrian Formation of Gondwana[J].Geology,1993,21:889-892.
[62] L I Z X,ZHANGL,POWELL CMCA.South China in Rodinia:Part of the missing link between Australia-East Antarctic and Lanrentia[J]?Geology,1995,23:407-410.[63] L I Z X.New paleomagnetic results from the“cap dolomite”of the Neoproterozoic Walsh Tittite,northwestern Australia[J].Precambrian Research,2000,100:359-370.
[64] 刘新秒.新元古代 Rodinia超大陆的研究进展[J].前寒武纪研究进展,2001,24(6):116-122.
[65] 王鸿祯,刘本培.地史学教程[M].北京:地质出版社,1978.70-103.
[66] MACOU IN M,BESSE J,ADER M et al. Combined paleomagnetic and isotopic data from Doushantuo carbonate,South China:implications for the“snowball Earth”hypothesis[J].Earth and Planetary Science Letters,2004,224:387-398.
[67] 陈长胜.海洋生态系统动力学与模型[M].北京:高等教育出版社,2003.1-287.
[68] 王约,王训练.黔东北新元古代陡山沱期宏体藻类的固着器特征及其沉积环境意义[J].微体古生物学报,2006,23(2):154-164.
[69] 胡南方.贵州震旦系陡山沱组生油岩特征[J].贵州地质,1997,14(3):244-251.
[70] 王约,王训练,高金汉,等.川北青川地区下寒武统沥青质砾石的发现及其意义[J].地质通报,2008,27(1):149-152.
Development of the Ed iacaran(Sin ian)multicellular organis ms and formation of the source rocks in the Yangtze area
WANG Yue1,LEILing-fang1,CHEN Hong-de2,HOU Ming-cai2
(1.School of Resources and Environm ent,Guizhou University,Guiyang550003,Guizhou,China;2.Institute of Sed im entary Geology,Chengdu University of Technology,Chengdu610059,Sichuan,China)
The productivity of organis ms and abundances of preserved organic matter are believed to be important conditions for the formation of source rocks.The multicellular organis ms appeared in substantial amounts in the Yangtze shallow sea during the Doushantuoan(Ediacaran).The black mudstones(shales)were once deposited as the water-rich gruel sed iments during the Doushantuoan.At that time,the oxidation-reduction interfaces were relatively shallower.The organic remains could be rapidly covered by the gruel sediments so as to form the source rocks.The Doushantuoan(Ediacaran)went through sharp changes in chemical,climatic and environmental aspects,and thusmay be a key period for the life evolution,development of organic productivity and for mation of the source rocks in the Yangtze area.
source rock;organic matter;oxygen;multicellular organis m;Doushantuoan;Yangtze plate
1009-3850(2010)03-0030-09
2010-02-15;改回日期2010-08-05
王约 (1964—),教授,主要从事古生物地层及沉积环境的研究。E-mail:gzyuewang@126.com
国家自然科学 (No.40739901)、贵州省省长基金和贵州省科技基金 (J-2010-2030)联合资助
TE122.1+13
A
