海水养殖池塘浮游植物碳的计算机模拟*
2010-01-05苏跃朋田相利冯翠梅董双林
苏跃朋,马 甡,田相利,冯翠梅,董双林
(中国海洋大学海水养殖教育部重点实验室,山东青岛266003)
海水养殖池塘浮游植物碳的计算机模拟*
苏跃朋,马 甡,田相利,冯翠梅,董双林
(中国海洋大学海水养殖教育部重点实验室,山东青岛266003)
应用光学显微镜法对实验系统浮游植物进行了野外定量研究,测定了系统中浮游植物的种类组成和生物碳素变动,为模型有效性验证提供了野外数据;根据Ecopath能量平衡模型原理建立了海水养殖池塘浮游植物碳的计算机模型,初步确定了包括15个参数的浮游植物模块参数库,通过单因子分析法进行参数敏感性检验,确定了3个敏感性参数,并利用野外实际测定数据进行了模型验证。模拟结果表明本模型成功模拟了海水养殖系统浮游植物碳变动。在建模过程中,光限制因子与水温为影响浮游植物碳的主要内源因素,浮游动物捕食作用为外源影响因素。
养殖池塘;浮游植物;碳;模型;敏感性参数
在全球各大海域中,浮游植物碳与初级生产力是物质转换和能量流动的关键环节,是一切草食、肉食乃至腐食生物赖以生存的物质基础[1]。在生态系统动力学模拟研究中,浮游植物模块具有举足轻重的地位。欧洲北海ECOHAM模型[2]、胶州湾ERSEM模型[3]、渤海NPZD生化模型[4]等动力学模拟研究中,浮游植物均作为生物箱式模型的基础模块进行建模。
在海水养殖系统中,浮游植物与浮游动物、细菌和腐屑共同组成“微食物环”[5],可以有效利用和转化残饵、粪便以及溶解有机物等污染物形成次级生产力,是清洁生产的重要环节。同时,浮游植物与养殖容量[6]、水生系统富营养化[7]、非经典生物操纵下赤潮控制[8]等研究密切相关。目前对于浮游植物的研究多见于Redfield系数下氮、磷、硅、铁等限制对其生长的影响[9],基于“微食物环”的系统动力学研究尚不多见。翟雪梅等[10]曾建立经典对虾养殖池塘鞭毛藻和浮游硅藻子模型,遗憾的是因缺乏实测数据而无法进行模型有效性的系统验证。Ecopath能量平衡模型是动态描述生态系统的高效生态通道建模方法[11]。本文借鉴前人工作经验,根据Ecopath能量平衡模型原理建立了海水养殖系统浮游植物碳的计算机模型,并利用野外实际测定数据进行了模型验证和参数敏感性分析,为海水养殖系统“微食物环”的模拟研究以及清洁生产技术开发奠定了基础。
1 材料和方法
研究对象为人工增氧型高密度对虾养殖系统。实验池塘位于天津汉沽区,池塘长方形,面积为0.20 hm2,池深4 m,水深度为3.0~3.5 m,底质类型为泥砂型,养殖用水为高盐度卤水和地下淡水混合的卤淡水,盐度为13~25,养殖周期中采用仅添水不排水的封闭式养殖模式。放养中国明对虾(Fenneropenaeus chinensis)虾苗,放养密度为250尾/m2。野外实验时间为2004年6月5日~8月20日。每15 d用2.50 L的有机玻璃采水器提取表、中、底3层水样混合。然后取混合水样1.00 L,以10%的I2溶液固定,在室内静置48 h后,浓缩、定容至50 mL。分析时取均匀样品注入0.10 m L的浮游植物计数框,用视野计数法在400×显微镜下计数小型浮游植物(≤20μm),用全片计数法在100×显微镜下计数大型浮游植物(>20μm),计数时为了使误差减少到10%,每次所计浮游植物细胞数均达到400个以上[12]。
采用Epp ley等[13]细胞体积法将浮游植物细胞数量转化为碳含量来计算浮游植物的生物量,计算公式为:

其中,C为浮游植物细胞含碳量,单位为pg;V为浮游植物细胞体积,单位为μm3。浮游植物各种类的细胞体积的计算采用Hillebrand等[14]的几何体积拟合法。
此外,从原始水样中另取50 mL混合水样,0.45μm滤膜过滤,再以N,N-二甲基甲酰胺萃取滤膜后,用3色分光光度法(检出限1μg/L,相对误差±10%)测定叶绿素a值[15]。
野外实验周期内共采集到浮游植物32属。其中,硅藻门10属,甲藻门6属,绿藻门4属,金藻、裸藻和蓝藻等种属所占比例均在10%以下,各门所占整体比例见图1。实验期间,浮游植物优势种主要有硅藻类的根管藻属(Rhizosolenia)和小环藻属(Cyclotella),绿藻类的小球藻属(Chlorella),以及甲藻类的纵裂甲藻属(Plerom onas)和原甲藻属(Prorocentrum)。

图1 实验周期内养殖池塘浮游植物的种属组成Fig.1 Constitutes of phytop lankton species in shrimp culture pond during the experimental period
2 模型建立
2.1 模型结构与说明
Ecopath假设模型对象的生态系统中全部生物功能组是稳定的,系统中各功能组的能量输入与输出保持平衡[11]。浮游植物碳模型的生态功能组包括:浮游植物光合作用、呼吸、死亡、胞外分泌和被浮游动物捕食5项[16],表达方程如下:

光照、水温和营养盐依次为影响浮游植物光合作用的效应因子[17],光照作用尤为重要,根据养殖环境中高腐屑含量的特点,光限制函数采用Wolfe公式[18],并设置光限制最小参数值。其中光照主要为太阳辐射。本文中水表层太阳辐射(W/m2)为系统强迫函数,实验周期中每5d用照度计测定,采用线性内插法得到函数曲线。影响浮游植物生长的温度效应采用最大生长速率下的指数公式[19]。则表达方程如下:
(ⅰ)光合作用=G×α0×LL t×浮游植物生物量
式中:G:浮游植物生长(1/d),
α0:营养盐影响因子,
LL t:光限制函数(1/d)。
其中:G=umax×e(α1×T)

式中:μmax:浮游植物最大生长速率(1/d);
α1:浮游植物生长速率的温度系数(1/℃);
T:为水温(℃),为系统强迫函数;
L t:水表层太阳辐射(W/m2),为系统强迫函数;
Iref:可被浮游植物利用的最大光照强度(W/m2);
UZP:单位浮游动物量(mgC/m3),为文献[20]中的浮游动物模块;
UPP:单位浮游植物量(m gC/m3),为模拟结果代入;
δ:腐屑悬浮系数(%);
KI:水体中物质的最大浓度(mgC/m3)。
Ecopath模型中将自然死亡和被捕食项作为能量输出的必备单元[12]。浮游植物模型中光合自养过程还伴随有呼吸和胞外分泌等生物过程,其中呼吸作用可作为光合自养作用的可逆过程,呼吸速率与浮游植物的生长速率为正相关[21],呼吸作用与自然死亡项均采用温度效应下的指数公式表达[2]。浮游动物对浮游植物的捕食采用lvlev指数函数[22],为了避免对浮游植物的过度摄食,设置摄食阈值[23],用IF函数表示。胞外分泌为浮游植物细胞溶解有机物代谢过程,其分泌量与浮游植物成一定的比例关系,采用线性方程表示[3]。则表达方程如下:
(ⅱ)呼吸=α2×e(α3×T)×浮游植物生物量
式中:α2:0℃时浮游植物呼吸速率(1/d),
α3:浮游植物呼吸温度系数(1/℃)。(ⅲ)死亡=α4×e(α5×T)×浮游植物生物量
经过与企业联系,结合多年教学经验,我们编写了更加符合旅游管理专业学生需要的校本合作教材《经济应用文写作教程》。该教材从教学与实践、实用与适用的角度出发,在基本理论讲清的同时,大量引入企业的实际写作案例,做到学生通过学习就可以雷同去进行工作实践,力争体现当前高职高专教学改革方针。编写组成员将充分吸收以往高职高专《应用文写作》课程教材的优点,以知识通俗化、应用化为原则,侧重将旅游管理专业的应用文文种写入教材;选择格式规范的导游词、解说词等案例、难易适度的课后习题及章后综合练习题,培养学生解决实际问题的能力。
式中:α4:0℃时浮游植物自然死亡速率(1/d),
α5:浮游植物自然死亡温度系数(1/℃)。
(ⅳ)胞外分泌=α6×光合作用
式中:α6:浮游植物溶出系数(1/d)。

式中:Gmax:0℃时浮游动物对浮游植物的最大捕食
率(1/d),
α7:浮游动物捕食浮游植物的温度系数
(1/℃),
PP0:饵料阈值浓度(mgC/m3),
λ:为lv lev常数。
浮游植物碳模型的能流关系模式图如图2所示。
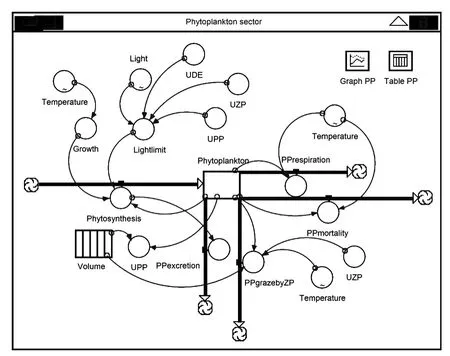
图2 浮游植物碳模型的能流关系模式图Fig.2 Concep tualmodel for phytop lankton carbon secto r showing pathways and linkages between different components
2.2 模型参数
合理选择和确定模型参数是数学模型构建中至关重要的环节[24]。模型参数的可靠性对于模型模拟结果的合理性、准确性等起着关键作用[25]。对于重要参数最好在实验现场进行测定,模型基本参数则通过文献查取适宜范围后进行选择和确定。本文所用参数见表1,除δ值采用沉积物罐收集法现场测定外,其他参照文献[10]、[22]和[27]。
3 模拟结果
本文模型是在STELLA○R5.1.1(High Performance System s,Inc.,Hanover,New Hampshire,USA)软件环境下开发运行,模型积分采用Euler’s方法,以1 d为时间步长进行数学模拟。该模型模拟了养殖周期90 d的浮游植物碳变动,以养殖0 d作为运行初始值,采用现场测定浮游植物碳和叶绿素a数据进行模型验证。结果(见图3)表明浮游植物碳模拟结果与实际测定值、叶绿素a吻合良好,其中浮游植物碳模拟值和实测值相关性极显著(n=5,r=0.95,P<0.01)。本模型成功模拟了海水养殖系统浮游植物碳变动。

表1 模型建立过程所需参数和取值Table 1 Parameters and its values in the process of modelmaking

图3 浮游植物碳测定和模拟结果Fig.3 Comparison of simulated results and determined data of phytoplankton carbon in water
浮游植物碳的同化过程主要为光合作用过程,而光合作用取决于光辐射的光强度[17],所以光限制因子在高密度精养池塘系统中具有重要作用。因为该养殖系统的高养殖生物量和高机械增氧量带来养殖水体中极高的悬浮腐屑负载量[20],极大程度地影响了浮游植物的光合作用过程,是造成养殖中后期浮游植物碳量下降的主要原因。在生态建模过程中,对光合作用的公式选取大多采用与最适光强、水深有关的Steele函数[27],而本文针对养殖水体高腐屑量的特点,采用与颗粒物浓度相关的Wolfe公式[18],有力地促进了模型的成功构建。此外,本文研究系统中硅藻所占比例高于30%,显著高于甲藻、绿藻、金藻和蓝藻等池塘常见种类(见图1)。研究期间,浮游植物优势种主要为硅藻类的根管藻属(Rhizosolenia)和小环藻属(Cyclotella)等。相对于其他藻类,浮游硅藻生活环境只需要很少的光,底栖硅藻可以生活在弱光照的深水处[28]。则藻类种属分析结果表明本研究环境仅适宜暗光生存的硅藻类,光限制因子除影响碳素同化过程外,对浮游植物的种属组成也具有重要影响作用。与光照因子具有相似性,温度对藻类结构也有着重要影响作用。大量硅藻种类为狭冷性种,适宜在春天和秋天生长繁殖,而蓝藻、绿藻和甲藻为广温性种,常在夏天水温较高时出现[29]。本研究系统在养殖中后期水温升高,藻类结构体系发生了从低温种类向广温种类演变,而适宜高水温生存的藻种又不适宜高浊度水体的光限制环境,是造成养殖中后期浮游植物碳量下降的又一重要因素。
相对于光照和温度而言,浮游植物生长对氮、磷等营养盐也有着基本需求,一般认为浮游植物以16∶1的摩尔比吸收氮磷,水环境中浮游植物的种群变动与氮磷比有一定的关系[9]。不同种类的浮游植物对氮、磷的最适浓度要求也有所不同,海水浮游植物要求氮、磷下限分别为无机氮1μmol/L,无机磷0.1μmol/L[30]。本研究系统中,氮磷比变动范围在8.8~30.9之间,无机N和无机P的浓度分别高于22和2.5μmol/L[20],远大于浮游植物对营养盐的下限要求。所以,在本实验系统中,营养盐浓度并不构成显著影响浮游植物生长和繁殖的限制因子。
影响浮游植物碳的外在因素主要为浮游动物的捕食作用。依照Harvery[31]的捕食假说,浮游植物和浮游动物二者在空间分布上呈相反的关系,虽然大型浮游动物对浮游植物的摄食作用受很多因素的影响,诸如水温、光照、浮游动物丰度、浮游植物丰度和有机碎屑的数量等,大多数学者仍然认为浮游动物是浮游植物生物量操纵的最主要因素[32-33]。本实验前期,浮游植物碳模拟值下降的主要原因即为浮游动物的摄食作用。此外,本实验研究系统的浮游动物以大中型(>200μm)个体为优势种类,包括:拟哲水蚤(Paraca lanus)、剑水蚤(Cyclops)和臂尾轮虫(B rachionus)等[20]。中国明对虾幼虾期可对植食性大型浮游动物具有显著捕食作用[34],所以在养殖前期放养对虾苗种后,浮游动物因虾苗摄食作用而显著下降[20],进而浮游动物对浮游植物碳的摄食作用随之降低,从而造成了养殖期25 d浮游植物的生长峰值(见图2)。可以看到在计算机模拟前期中,浮游动物与浮游植物具有此消彼长的现象,卢静等[33]在传统养虾池塘的研究中也观察到了相同的现象。同时发现,浮游植物碳随着浮游动物数量变动而出现下降和上升峰值的模拟结果,因恰好出现在采样测定间期而并未被实际测定结果观察到。该结果表明,对于复杂多变的水生生态系统而言,计算机模拟可以灵敏、准确地反映出养殖系统内部变化规律。通过模型的预测分析功能,可进行健康高效养殖关键技术的筛选,从而为养殖生产服务。
4 模型敏感性分析
灵敏度分析(Sensitivity analysis)在系统分析中有很重要的作用,其目的在于确定模型中哪些方面最容易在系统描述中引进不确定性,具有重要的生态学意义[35]。
本文采用单因子变化法,在模型标准运行状态下改变特定参数的值,取状态变量在各个时间点的相对变化量的平均值,除以参数的相对变化,即得出状态变量的敏感度[36]。对于参数变化范围采用文献[37]的原则,即在参数估计的范围内,将预分析的参数增加10%或减少10%。对模型中的主要参数进行了敏感度分析(见表2)。以敏感度>1的为敏感参数,1~0.5之间为较敏感参数,<0.5为不敏感参数。

表2 浮游植物模型参数敏感性分析Table 2 Sensitivity analysis in phytop lankton model
单因子参数分析表明,浮游植物最大生长速率、营养盐影响因子和0℃时浮游动物对浮游植物的最大捕食率均为敏感性参数;可被浮游植物利用的最大光照强度和水表层太阳辐射为较敏感参数。表明与敏感性参数相关的不同水环境中浮游植物种类、营养盐富营养化程度和浮游动物生物量以及种类组成都可以显著影响浮游植物碳量。
5 结语
本文根据Ecopath能量平衡模型原理建立了海水养殖池塘浮游植物碳的计算机模型,浮游植物碳模型的生态功能组包括浮游植物光合作用、呼吸、死亡、胞外分泌和被浮游动物捕食5项。浮游植物碳的模拟结果与实际测定值值相关性极显著,本模型成功模拟了海水养殖池塘浮游植物碳变动。在建模过程中,光限制因子与水温为影响浮游植物碳的主要内源因素,浮游动物捕食作用为外源影响因素。
参数分析表明,浮游植物最大生长速率、营养盐影响因子和浮游动物对浮游植物的最大捕食率均为敏感性参数。则在特定养殖水域的建模过程中,需进行养殖环境优势藻种类鉴定,优势种生长速度测定,以及养殖环境中浮游动物优势种对特定浮游植物的摄食强度率定,进而利用本模型完成计算机模拟预测,可以大大提高对特定养殖水域浮游植物碳模拟的精度。
[1] 刘慧,董双林,方建光.全球海域营养盐限制研究进展[J].海洋科学,2002,8:47-53.
[2] Moll A.Regional distribution of p rimary p roduction in the No rth Sea simulated by a three-dimensionalmodel[J].Journal of Marine Systems,1998,16:151-170.
[3] 俞光耀,吴增茂,张志南.胶州湾北部水层生态动力学模型与模拟I.胶州湾北部水层生态动力学模型[J].青岛海洋大学学报,1999,29(3):421-428.
[4] 高会旺,王强.1999年渤海浮游植物生物量的数值模拟[J].中国海洋大学学报:自然科学版,2004,34(5):867-873.
[5] Azam F,Fenchel T,Field J G,et al.The ecological role of water column microbes in the sea[J].Mar Ecol Prog Ser,1983,10:257-263.
[6] 李长松,房斌,王慧,等.贝类养殖容量研究进展[J].上海水产大学学报,2007,16(5):478-483.
[7] 张传松,王修林,朱德弟,等.营养盐在东海春季大规模赤潮形成过程中的作用[J].中国海洋大学学报:自然科学版,2007,37(6):1002-1006.
[8] 张根芳,邓闽中,方爱萍.蚌、鱼养殖模式对水体富营养化控制作用的研究[J].中国海洋大学学报:自然科学版,2005,35(3):491-495.
[9] 彭欣,宁修仁,蔡岦明,等.浮游植物生长上行效应的研究进展[J].海洋学研究,2006,24(3):64-76.
[10] 翟雪梅,张志南.虾池生态系浮游生物亚模型[J].青岛海洋大学学报,1999,21(1):94-100.
[11] 苏纪兰,唐启升.中国海洋生态系统动力学研究:Ⅱ渤海生态系统的动力学过程[M].北京:科学出版社,2002:289-305.
[12] Lund,J W,Kipling C,LeCren E D.The inverted microscope method of estimating algal numbersand the statistical basisof estimations by counting[J].Hydrobiological,1958,11:143-170.
[13] Eppley R W,Reid F M,Strickland J D.The ecology of the plankton of La Jolla California,in the period Ap ril through September 1967[C].∥Stickland,J D(Ed.),Estimates of Phytoplankton Crop Size,Grow th Rate and Primary Production PartⅢ,Bull Scripps Inst Oceanogr,1970,17:33-42.
[14] Hillebrand H,Durselen C D,Kirschtel D,et al.Biovolume calculation for pelagic and benthic microalgae[J].J Phycol,1999,35:401-424.
[15] Jeffrey SW,Humphrey G F.New spectrophotometric equations for determining chlorophylls a,b,and c in higher plants,algae and natural phytoplankton[J].Biochen Physiol Pflanzen,1975,167:191-194.
[16] Varela R A,Cruzado A,Gabaldon J E.Modelling primary production in the North Sea using the European Regional Seas ecosystem model[J].Netherlands Journal of Sea Research,1995,33(3/4):337-361.
[17] 杨东方,陈生涛,胡均,等.光照、水温和营养盐对浮游植物生长重要影响大小的顺序[J].海洋环境科学,2007,26(3):201-207.
[18] Wolfe,J R,Zweig,R D,Engstrom,D G.A computer simulation model of solar-algae pond ecosystem[J].Ecological Modelling,1986,34:1-59.
[19] Epply R W.Temperature and phytoplankton grow th in the sea[J].Fish Bulletin U.S.,1972,70:1063-1085.
[20] 苏跃朋.中国明对虾精养池塘生态系统及动力学模型的研究[D].青岛:中国海洋大学,2006.
[21] Law s E,Caperon J.Carbon and nitrogen metabolism by Monochrysis lutheri:measurement of grow th rate dependent respiration rates[J].Marine Biology,1976,36(1):85-97.
[22] Franks P J,Wroblew ski J S,Flierl G R.Behavior of a simple plankton model with food-level acclimation by herbivores[J].Marine Biology,1986,91:121-129.
[23] Radach G,Moll A,Willebrand J R.Estimation of the variability of production by simulating annual cycles of phytoplankton in the central North Sea[J].Progress in Oceanography,1993,31(4):339-419.
[24] Harmon R,Challenor P A.Markov chain Monte Carlo method for estimation and assimilation into models[J].Ecological Modelling,1997,101(1):41-59.
[25] Schartau M,Oschlies A,Willebrand J R.Parameter estimatesof a zero-dimensional ecosystem model applying the adjoint method[J].Deep-Sea ResearchⅡ,2001,48:1769-1800.
[26] 石晓勇.长江口营养盐、石油烃对海洋生态系统影响及动力学研究[D].青岛:中国海洋大学,2004.
[27] Steele J H.Environmental control pf photosynthesis in the sea[J].Limnology and Oceanography,1962,7:137-150.
[28] 福迪.藻类学[M].上海:上海科学技术出版社,1980:215.
[29] Whitford L A,Schumacher GJ.Effect of current on mineral uptake and respiration by fresh-water alga[J].Limnology and Oceanography,1961,6:423-425.
[30] Justic D,Rabalais N,Turner R,et al.Changes in nutrient structure of river-dominated coastal waters:stoichio-metric nutrient balance and its consequences[J].Estuarine,Coastal and Shelf Science,1995,40:339-356.
[31] Harvey H W.Recent advances in the chemistry and biology of sea water[J].The Quarterly Review of Biology,1946,21(3):275-276.
[32] 陈济丁,任久长,蔡晓明.利用大型浮游动物控制浮游植物过量生长的研究[J].北京大学学报:自然科学版,1996,31(3):373-381.
[33] 卢静,李德尚,董双林.对虾池的放养密度对浮游生物群落的影响[J].水产学报,2000,24(3):240-246.
[34] 孙修涛,李健,麻次松,等.中国对虾虾池几种动物的摄食能力和选择性研究[J].海洋科学,1995,3:1-4.
[35] 徐崇刚,胡远满,常禹,等.生态模型的灵敏度分析[J].应用生态学报,2004,15(6):1056-1062.
[36] Battaglia M,Sands P.Aplication of sensitivity analysis to amodel of Eucalyptus globulus plantation p roductivity[J].Ecological Modelling,1998,111:237-259.
[37] Jφgensen S E.Fundamentals of Ecological Modelling(2nd Edition)[M].Elsevier:Amsterdam,1994:630.
Computer Simulation of the Phytoplankton Carbon in Mariculture Pond
SU Yue-Peng,MA Shen,TIAN Xiang-Li,FENG Cui-M ei,DONG Shuang-Lin
(The Key Labo rato ry of Maricultere.M inistry of Education,Ocean University of China,Qingdao 266003,China)
The species and the carbon of phytop lankton are studied in the intensive shrimp culture ecosystem with the use of the optical microscope,for provided the field data to the validation of the dynamical model of phytoplankton carbon.Acomputermodel is built up according to the Ecopath model method.A module stock which consisted of 15 parameters is primarily established,and 3 sensitivity parameters are found by sensitivity analysis.The results show that the simulated data are in accordance with the field data of phytoplankton carbon well.In simulation,the factors of light limit and water temperature are the interior facto rs of model building,and the predation of zoop lankton is the exterior ones.
mariculture pond;phytop lankton;carbon;model;sensitivity parameter
S917
A
1672-5174(2010)12-037-07
国家科技支撑计划项目(2007BAD43B06);国家高技术研究发展计划项目(2002AA 648010)资助
2009-02-22;
2009-04-08
苏跃朋(1978-),男,博士,讲师。Tel:0532-82032195;E-mail:suyuepeng@ouc.edu.cn
责任编辑 于 卫
